
Does the Presence of Shade Trees and Distance to the Forest Affect Detection Rates of Terrestrial Vertebrates in Coffee Home Gardens?

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Data Collection
2.3. Data Analysis
3. Results
3.1. Detection Rates
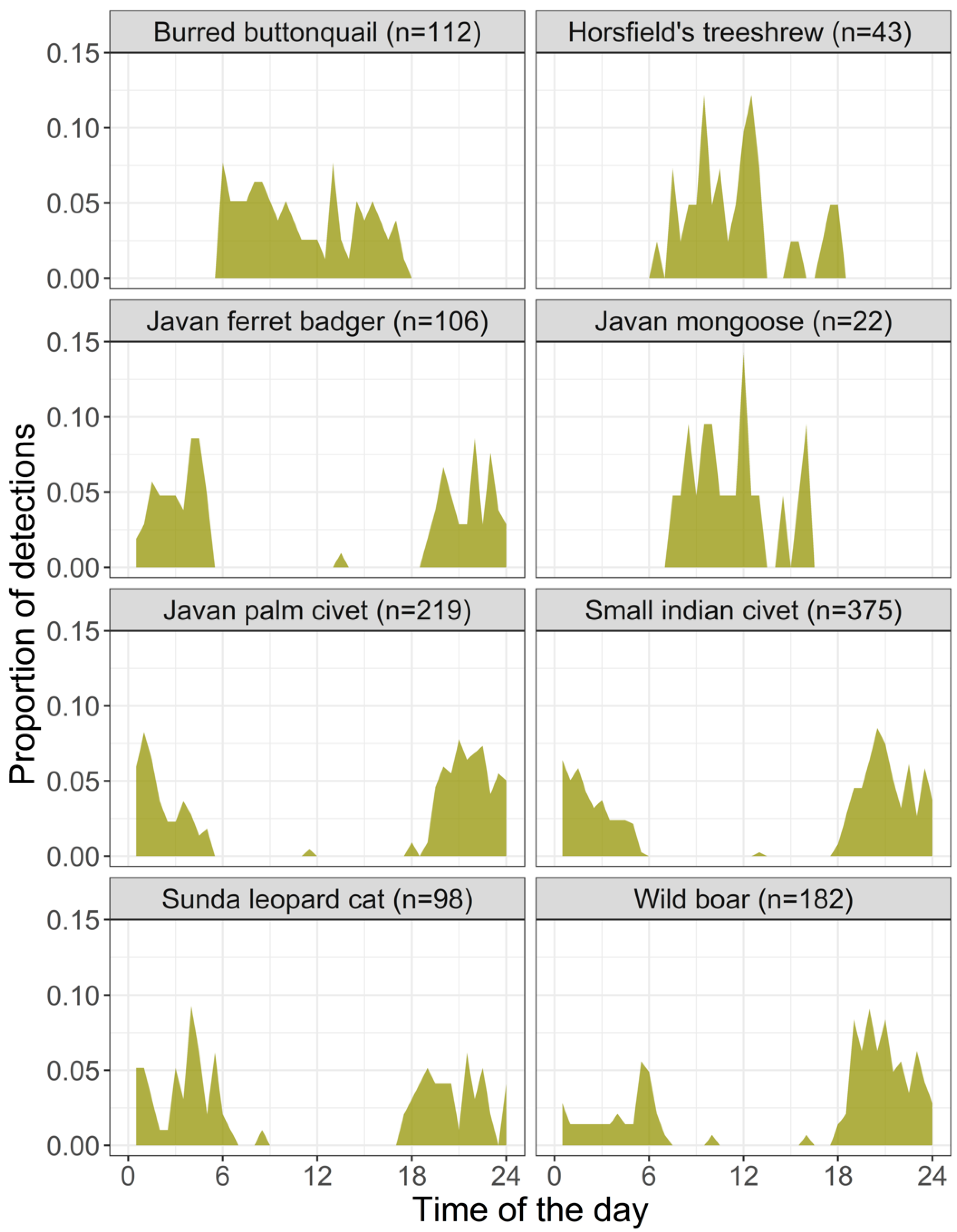
3.2. Spatial and Temporal Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermier, C.G.; de Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Laurance, W.F.; Sayer, J.; Cassman, K.G. Agricultural expansion and its impacts on tropical nature. Trends Ecol. Evol. 2014, 29, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Laurance, W.F. Have we overstated the tropical biodiversity crisis? Trends Ecol. Evol. 2007, 22, 65–70. [Google Scholar] [CrossRef]
- Dobrovolski, R.; Diniz-Filho, J.A.F.; Loyola, R.D.; Júnior, P.D.M. Agricultural expansion and the fate of global conservation priorities. Biodivers. Conserv. 2011, 20, 2445–2459. [Google Scholar] [CrossRef]
- Fischer, J.; Brosi, B.; Daily, G.C.; Ehrlich, P.R.; Goldman, R.; Goldstein, J.; Lindenmayer, D.B.; Manning, A.D.; Mooney, A.H.; Pejchar, L.; et al. Should agricultural policies encourage land sparing or wildlife-friendly farming? Front. Ecol. Environ. 2008, 6, 380–385. [Google Scholar] [CrossRef]
- Tscharntke, T.; Clough, Y.; Wanger, T.C.; Jackson, L.; Motzke, I.; Perfecto, I.; VanderMeer, J.; Whitbread, A. Global food security, biodiversity conservation and the future of agricultural intensification. Biol. Conserv. 2012, 151, 53–59. [Google Scholar] [CrossRef]
- Phalan, B. What have we learned from the land sparing-sharing model? Sustainability 2018, 10, 1760. [Google Scholar] [CrossRef] [Green Version]
- Bhagwat, S.A.; Willis, K.J.; Birks, H.J.B.; Whittaker, R.J. Agroforestry: A refuge for tropical biodiversity? Trends Ecol. Evol. 2008, 23, 261–267. [Google Scholar] [CrossRef]
- Santos-Heredia, C.; Andresen, E.; Zàtare, D.A.; Escobar, F. Dung beetles and their ecological functions in three agroforestry systems in the Lacandona rainforest of Mexico. Biodivers. Conserv. 2018, 27, 2379–2394. [Google Scholar] [CrossRef]
- Jha, S.; Bacon, C.M.; Philpott, S.M.; Méndez, V.E.; Läderach, P.; Rice, R.A. Shade coffee: Update on a disappearing refuge for biodiversity. BioScience 2014, 64, 416–428. [Google Scholar] [CrossRef] [Green Version]
- Tscharntke, T.; Clough, Y.; Bhagwat, S.A.; Buchori, D.; Faust, H.; Hertel, D.; Hölscher, D.; Juhrbandt, J.; Kessler, M.; Perfecto, I.; et al. Multifunctional shade-tree management in tropical agroforestry landscapes—A review. J. Appl. Ecol. 2011, 48, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, R.; Bichier, P.; Angon, A.C.; Reitsma, R. Bird populations in shade and sun coffee plantations in Central Guatemala. Conserv. Biol. 1997, 11, 448–459. [Google Scholar] [CrossRef]
- Muriel, S.B.; Kattan, G.H. Effects of patch size and type of coffee matrix on ithomiine butterfly diversity and dispersal in cloud-forest fragments. Conserv. Biol. 2009, 23, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Jha, S.; Bacon, C.M.; Philpott, S.M.; Rice, R.A.; Méndez, V.E.; Läderach, P.; Campbell, W.B.; López Ortiz, S. A review of ecosystem services, farmer livelihoods, and value chains in shade coffee agroecosystems. In Integrating Agriculture, Conservation and Ecotourism: Examples from the Field; Campbell, W., Lopez Ortiz, S., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 141–208. [Google Scholar]
- Perfecto, I.; Rice, R.A.; Greenberg, R.; VanderVoort, M.E. Shade coffee: A disappearing refuge for biodiversity. BioScience 1996, 46, 598–608. [Google Scholar] [CrossRef] [Green Version]
- Caravaggi, A.; Banks, P.B.; Burton, A.C.; Finlay, C.M.V.; Haswell, P.M.; Hayward, M.W.; Rowcliffe, M.J.; Wood, M.D. A review of camera trapping for conservation behaviour research. Remote. Sens. Ecol. Conserv. 2017, 3, 109–122. [Google Scholar] [CrossRef]
- Glen, A.S.; Cockburn, S.; Nichols, M.; Ekanayake, J.; Warburton, B. Optimising camera traps for monitoring small mammals. PLoS ONE 2013, 8, e67940. [Google Scholar] [CrossRef]
- Debata, S.; Swain, K.K. Estimating mammalian diversity and relative abundance using camera traps in a tropical deciduous forest of Kuldiha Wildlife Sanctuary, eastern India. Mammal. Study 2018, 43, 45–53. [Google Scholar] [CrossRef]
- Rovero, F.; Marshall, A.R. Camera trapping photographic rate as an index of density in forest ungulates. J. Appl. Ecol. 2009, 46, 1011–1017. [Google Scholar] [CrossRef] [Green Version]
- Rovero, F.; Martin, E.; Rosa, M.; Ahumada, J.A.; Spitale, D. Estimating species richness and modelling habitat preferences of tropical forest mammals from camera trap data. PLoS ONE 2014, 9, e103300. [Google Scholar] [CrossRef]
- Rode-Margono, E.J.; Voskamp, A.; Spaan, D.; Lehtinen, J.K.; Roberts, P.D.; Nijman, V.; Nekaris, K.A.I. Records of small carnivores and of medium-sized nocturnal mammals on Java, Indonesia. Small Carniv. Conserv. 2014, 50, 1–11. [Google Scholar]
- Top Coffee Producing Countries. WorldAtlas. Available online: www.worldatlas.com/articles/top-coffee-producing-countries.html (accessed on 10 June 2021).
- von Rintelen, K.; Arida, E.; Häuser, C. A review of biodiversity-related issues and challenges in megadiverse Indonesia and other Southeast Asian countries. Res. Ideas Outcomes 2017, 3, e20860. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.; Barton, D.; Johnson, M.D.; Wendt, C.; Milligan, M.C.; Njoroge, P.; Gichuki, P. Bird communities in sun and shade coffee farms in Kenya. Glob. Ecol. Conserv. 2015, 4, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.S.; Peres, C.A.; Bogoni, J.A.; Cassano, C.R. Use of agroecosystem matrix habitats by mammalian carnivores (Carnivora): A global-scale analysis. Mamm. Rev. 2018, 48, 312–327. [Google Scholar] [CrossRef]
- Cheyne, S.; Sastramidjaja, W.J.; Muhalir; Rayadin, Y.; Macdonald, D.W. Mammalian communities as indicators of disturbance across Indonesian Borneo. Glob. Ecol. Conserv. 2016, 7, 157–173. [Google Scholar] [CrossRef] [Green Version]
- Campera, M.; Budiadi, B.; Adinda, E.; Ahmad, N.; Balestri, M.; Hedger, K.; Imron, M.A.; Manson, S.; Nijman, V.; Nekaris, K.A.I. Fostering a wildlife-friendly program for sustainable coffee farming: The case of small-holder farmers in Indonesia. Land 2021, 10, 121. [Google Scholar] [CrossRef]
- Nekaris, K.A.I.; Poindexter, S.; Reinhardt, K.D.; Sigaud, M.; Cabana, F.; Wirdateti, W.; Nijman, V. Coexistence between Javan slow lorises (Nycticebus javanicus) and humans in a dynamic agroforestry landscape in West Java, Indonesia. Int. J. Primatol. 2017, 38, 303–320. [Google Scholar] [CrossRef] [Green Version]
- Cusack, J.J.; Dickman, A.J.; Kalyahe, M.; Rowcliffe, J.M.; Carbone, C.; Macdonald, D.W.; Coulson, T. Revealing kleptoparasitic and predatory tendencies in an African mammal community using camera traps: A comparison of spatiotemporal approaches. Oikos 2017, 126, 812–822. [Google Scholar] [CrossRef]
- Birot, H.; Campera, M.; Imron, M.A.; Nekaris, K.A.I. Artificial canopy bridges improve connectivity in fragmented landscapes: The case of Javan slow lorises in an agroforest environment. Am. J. Primatol. 2019, 82, e23076. [Google Scholar] [CrossRef]
- Wood, S.N. Package ‘mgcv’. pdf. R Package Version. 2018. Available online: http://cran.r-project.org/web/packages/mgcv/mgcv (accessed on 1 June 2021).
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall: Boca Raton, FL, USA, 2017. [Google Scholar]
- Wei, T.; Simko, V. R Package “corrplot”: Visualization of a Correlation Matrix. 2021. Available online: https://cran.r-project.org/web//packages/corrplot/corrplot (accessed on 1 June 2021).
- Luskin, M.; Christina, E.; Kelley, L.; Potts, M. Modern hunting practices and wild meat trade in the oil palm plantation-dominated landscapes of Sumatra, Indonesia. Hum. Ecol. 2014, 42, 35–45. [Google Scholar] [CrossRef]
- Marsh, C.; Nekaris, K.A.I.; Wirdateti, W. Crop raiding by Sus scrofa leads to pig fighting arenas in West Java. Suiform Sound. 2016, 14, 21–23. [Google Scholar]
- Sugiyarto; Nayasilana, I.N.; Aditya. The suburban forest as a habitat of eagles (Accipitridae): A case study in Gunung Bromo University Forest, Karanganyar, Central Java, Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2020, 590, 012007. [Google Scholar] [CrossRef]
- Irawan, N.; Pudyatmoko, S.; Yuwono, P.S.H.; Tafrichan, M.; Giordano, A.J.; Imron, M.A. The importance of unprotected areas as habitat for the leopard cat (Prionailurus bengalensis javanensis Desmarest, 1816) on Java, Indonesia. J. Ilmu Kehutan. 2020, 14, 198–212. [Google Scholar]
- Grassman, L.I., Jr.; Tewes, M.E.; Silvy, N.J.; Kreetiyutanont, K. Spatial organization and diet of the leopard cat (Prionailurus bengalensis) in north-central Thailand. J. Zool. 2005, 266, 45–54. [Google Scholar] [CrossRef]
- Rajaratnam, R.; Sunquist, M.; Rajaratnam, L.; Ambu, L. Diet and habitat selection of the leopard cat (Prionailurus bengalensis borneoensis) in an agricultural landscape in Sabah, Malaysian Borneo. J. Trop. Ecol. 2007, 23, 209–217. [Google Scholar] [CrossRef]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape perspectives on agricultural intensification and biodiversity—ecosystem service management. Ecol. Lett. 2004, 8, 857–874. [Google Scholar] [CrossRef]
- Cassano, C.R.; Barlow, J.; Pardini, R. Large mammals in an agroforestry mosaic in the Brazilian Atlantic forest. Biotropica 2012, 44, 818–825. [Google Scholar] [CrossRef]
- Wibisono, H.T.; Wahyudi, H.A.; Wilianto, E.; Pinondang, I.M.R.; Primajati, M.; Liswanto, D.; Linkie, M. Identifying priority conservation landscapes and actions for the Critically Endangered Javan leopard in Indonesia: Conserving the last large carnivore in Java Island. PLoS ONE 2018, 13, e0198369. [Google Scholar] [CrossRef]
- Balme, G.A.; Slotow, R.; Hunter, L.T.B. Impact of conservation interventions on the dynamics and persistence of a persecuted leopard (Panthera pardus) population. Biol Conserv. 2009, 142, 2681–2690. [Google Scholar] [CrossRef]
- Nijman, V. Group composition and monandry in grizzled langurs, Presbytis comata, on Java. Folia Primatol. 2017, 88, 237–254. [Google Scholar] [CrossRef] [Green Version]
- Setiawan, A.; Wibisono, Y.; Nugroho, T.S.; Agustin, I.Y.; Imron, M.A.; Pudyatmoko, S.; Djuwantoko. Javan surili: A survey population and distribution in Mt. Slamet Central Java, Indonesia. J. Primatol. Indones. 2010, 7, 51–54. [Google Scholar]
- Santosa, Y.; Rahmawati, C.; Utami, C.Y. Estimation of demographic parameters, spatial distribution of activity and its habitat types used of Javan Surili (Presbytis comata) in Sigedong Forest Block, Ciremai Mount National Park. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2020; Volume 528. [Google Scholar] [CrossRef]
- Nakamoto, A.; Kinjo, K.; Baba, M.; Doi, T.; Izawa, M. Mammalian fauna in a coconut palm plantation recorded by photo-traps and sightings in West Java, Indonesia. Bull. Kitakyushu Mus. Nat. Hist. Hum. Hist. Ser. A (Nat. Hist.) 2006, 4, 121–123. [Google Scholar]
- Husodo, T.; Febrianto, P.; Megantara, E.N.; Shanida, S.S.; Pujianto, M.P. Diversity of mammals in forest patches of Cisokan, Cianjur, West Java, Indonesia. Biodiversitas 2019, 20, 1281–1288. [Google Scholar] [CrossRef] [Green Version]
- Mustikasari, I.A.; Withaningsih, S.; Megantara, E.N.; Husodo, T.; Parikesit, P. Population and distribution of Sunda porcupine (Hystrix javanica F. Cuvier, 1823) in designated area of Cisokan Hydropower, West Java, Indonesia. Biodiversitas 2019, 20, 762–769. [Google Scholar] [CrossRef] [Green Version]
- Ario, A. Preliminary study on bird and mammal diversity at ecosystem restored areas in the Gunung Gede Pangrango National Park, West Java, Indonesia. Indones. J. Appl. Environ. Stud. 2020, 1, 34–42. [Google Scholar]
- Moore, J.H.; Sittimongkol, S.; Campos-Arceiz, A.; Sumpah, T.; Eichhorn, M.P. Fruit gardens enhance mammal diversity and biomass in a southeast Asian rainforest. Biol. Conserv. 2016, 194, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhang, J.; Slade, E.; Zhang, L.; Palomares, F.; Chen, J.; Wang, X.; Zhang, S. Dietary shifts in relation to fruit availability among masked palm civets (Paguma larvata) in central China. J. Mammal. 2008, 89, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.A.D.S.; Alvarez, M.R.D.V.; Mariano-Neto, E.; Cassano, C.R. Is shadier better? The effect of agroforestry management on small mammal diversity. Biotropica 2020, 52, 470–479. [Google Scholar] [CrossRef]
- Nekaris, K.A.I.; Handby, V.; Campera, M.; Birot, H.; Hedger, K.; Eaton, J.; Imron, M.A. Implementing and monitoring the use of artificial canopy bridges by mammals and birds in an Indonesian agroforestry environment. Diversity 2020, 12, 399. [Google Scholar] [CrossRef]
- Campera, M.; Balestri, M.; Manson, S.; Hedger, K.; Ahmad, N.; Nijman, V.; Budiadi, B.; Imron, M.A.; Nekaris, K.A.I. Shade trees and agrochemical use affect butterfly assemblages in coffee home gardens. Agric. Ecosyst. Environ. 2021, 319, 107547. [Google Scholar] [CrossRef]
- Nekaris, K.A.I. The Little Fireface Project: Community conservation of Asia’s slow lorises via ecology, education, and empowerment. In Ethnoprimatology Developments in Primatology: Progress and Prospects; Weller, M., Ed.; Springer Science and Business Media: New York, NY, USA, 2016; pp. 259–272. [Google Scholar]
- Broadley, K.; Burton, A.C.; Avgar, T.; Boutin, S. Density-dependent space use affects interpretation of camera trap detection rates. Ecol. Evol. 2019, 9, 14031–14041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappelle, N.; Howe, E.J.; Boesch, C.; Kühl, H.S. Estimating animal abundance and effort–precision relationship with camera trap distance sampling. Ecosphere 2021, 12, e03299. [Google Scholar] [CrossRef]
- Howe, E.J.; Buckland, S.T.; Després-Einspenner, M.-L.; Kühl, H.S. Distance sampling with camera traps. Methods Ecol. Evol. 2017, 8, 1558–1565. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Detection Rate Per Coffee Home Garden (Ind./Day) | Detection Rate Forest (Ind./Day) | ||
|---|---|---|---|---|
| Mean | 95% CI | Mean | 95% CI | |
| Small Indian civet Viverricula indica | 0.0593 * | ±0.0400 | 0.0013 | ±0.0162 |
| Small rodents, unknown spp. | 0.0426 | ±0.0303 | 0.2654 * | ±0.1302 |
| Javan palm civet Paradoxurus musangus javanicus | 0.0293 | ±0.0172 | 0.0355 | ±0.0302 |
| Wild boar Sus scrofa | 0.0203 | ±0.0208 | 0.2451 * | ±0.1268 |
| Barred buttonquail Turnix suscitator | 0.0161 | ±0.0122 | 0.0051 | ±0.0003 |
| Javan ferret badger Melogale orientalis | 0.0150 | ±0.0125 | 0.0165 | ±0.1647 |
| Sunda leopard cat Prionailurus javanensis | 0.0141 | ±0.0098 | 0.0032 | ±0.0079 |
| Horsfield’s treeshrew Tupaia javanica | 0.0066 | ±0.0051 | 0.0159 | ±0.0071 |
| Javan mongoose Herpestes javanicus | 0.0025 | ±0.0025 | 0.0006 | ±0.0081 |
| Javan slow loris Nycticebus javanicus | 0.0028 | ±0.0034 | - | - |
| Yellow-throated marten Martes flavigula | 0.0002 | ±0.0003 | 0.0095 * | ±0.0086 |
| Sunda stink badger Mydaus javanensis | 0.0001 | ±0.0001 | - | - |
| Javan leopard Panthera pardus melas | - | - | 0.0038 | ±0.0164 |
| Grizzled langur Presbytis comata | - | - | 0.0032 | ±0.0079 |
| Sunda porcupine Hystrix javanica | - | - | 0.0013 | ±0.0160 |
| Response Variable | Predictor | Estimate | Std. Error | T | p | Smooth Term | p | |
|---|---|---|---|---|---|---|---|---|
| Edf | F | |||||||
| Barred buttonquail | Intercept | −4.205 | 0.886 | −4.7 | <0.001 * | |||
| Sun-exposed | −0.120 | 0.784 | −0.2 | 0.880 | ||||
| Detection distance | −0.008 | 0.116 | −0.1 | 0.949 | ||||
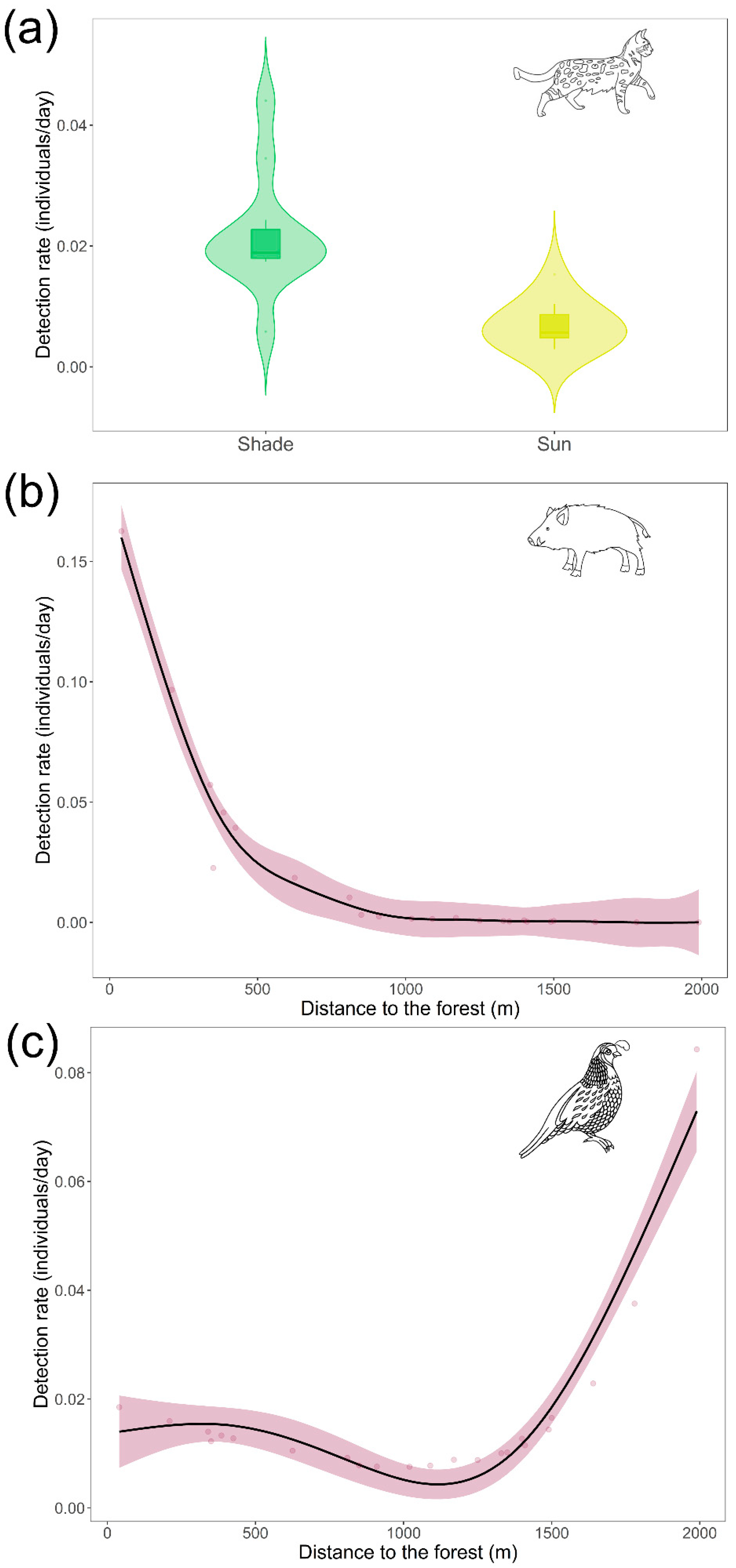
| (Distance to forest) | 2.213 | 3.7 | 0.046 * | |||||
| Horsfield’s treeshrew | Intercept | −5.036 | 1.006 | −5.0 | <0.001 * | |||
| Sun-exposed | −0.434 | 0.871 | −0.3 | 0.624 | ||||
| Detection distance | −0.062 | 0.122 | −0.5 | 0.618 | ||||
| (Distance to forest) | 2.613 | 1.3 | 0.312 | |||||
| Javan ferret badger | Intercept | −2.928 | 1.186 | −2.5 | 0.023 | |||
| Sun-exposed | −1.417 | 1.002 | −1.4 | 0.173 | ||||
| Detection distance | −0.126 | 0.196 | −0.6 | 0.528 | ||||
| (Distance to forest) | 1.000 | 1.7 | 0.206 | |||||
| Javan mongoose | Intercept | −4.561 | 2.623 | −1.7 | 0.099 | |||
| Sun-exposed | −0.563 | 1.339 | −0.4 | 0.679 | ||||
| Detection distance | −0.881 | 0.406 | −2.2 | 0.044 * | ||||
| (Distance to forest) | 1.942 | 0.9 | 0.479 | |||||
| Javan palm civet | Intercept | −2.991 | 0.931 | −3.2 | 0.005 * | |||
| Sun-exposed | −0.611 | 0.848 | −0.7 | 0.480 | ||||
| Detection distance | −0.061 | 0.131 | −0.5 | 0.647 | ||||
| (Distance to forest) | 1.000 | 0.5 | 0.508 | |||||
| Small rodents | Intercept | −1.994 | 1.278 | −1.6 | 0.135 | |||
| Sun-exposed | −0.596 | 0.925 | −0.6 | 0.527 | ||||
| Detection distance | −0.162 | 0.221 | −0.7 | 0.472 | ||||
| (Distance to forest) | 1.000 | 0.0 | 0.870 | |||||
| Small Indian civet | Intercept | −3.240 | 0.782 | −4.1 | <0.001 * | |||
| Sun-exposed | 0.435 | 0.645 | 0.7 | 0.508 | ||||
| Detection distance | −0.027 | 0.086 | −0.3 | 0.760 | ||||
| (Distance to forest) | 2.539 | 2.2 | 0.130 | |||||
| Sunda leopard cat | Intercept | −3.443 | 0.652 | −5.3 | <0.001 * | |||
| Sun-exposed | −1.785 | 0.805 | −2.2 | 0.039 * | ||||
| Detection distance | −0.095 | 0.126 | −0.8 | 0.461 | ||||
| (Distance to forest) | 1.450 | 1.0 | 0.295 | |||||
| Wild boar | Intercept | −5.167 | 1.671 | −3.1 | 0.006 * | |||
| Sun-exposed | −0.862 | 1.122 | −0.8 | 0.452 | ||||
| Detection distance | −0.063 | 0.243 | −0.3 | 0.797 | ||||
| (Distance to forest) | 1.000 | 8.8 | 0.008 * | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campera, M.; Hedger, K.; Birot, H.; Manson, S.; Balestri, M.; Budiadi, B.; Imron, M.A.; Nijman, V.; Nekaris, K.A.I. Does the Presence of Shade Trees and Distance to the Forest Affect Detection Rates of Terrestrial Vertebrates in Coffee Home Gardens? Sustainability 2021, 13, 8540. https://doi.org/10.3390/su13158540
Campera M, Hedger K, Birot H, Manson S, Balestri M, Budiadi B, Imron MA, Nijman V, Nekaris KAI. Does the Presence of Shade Trees and Distance to the Forest Affect Detection Rates of Terrestrial Vertebrates in Coffee Home Gardens? Sustainability. 2021; 13(15):8540. https://doi.org/10.3390/su13158540
Chicago/Turabian StyleCampera, Marco, Katherine Hedger, Hélène Birot, Sophie Manson, Michela Balestri, Budiadi Budiadi, Muhammad Ali Imron, Vincent Nijman, and K. A. I. Nekaris. 2021. "Does the Presence of Shade Trees and Distance to the Forest Affect Detection Rates of Terrestrial Vertebrates in Coffee Home Gardens?" Sustainability 13, no. 15: 8540. https://doi.org/10.3390/su13158540