
Abstract
Although human skin odor is thought to be the cue that anthropophilic mosquitoes use to discriminate us from other potential hosts, the precise details of how they use skin odor to find and land on a human is unclear. We found that Aedes aegypti land on a source of skin odor without a co-located visual cue. By collecting human odor on glass beads and using identical glass beads to visually conceal skin odor and heat cues, we were able to study mosquito landing on skin odor, heat, and visual cues separately. Landing is necessary for blood feeding which is a required behavior for the Aedes aegypti life cycle as well as the behavior responsible for the epidemiological impact of mosquitoes. Therefore, we consider it to be the diagnostic measure of the importance of a host cue. In two-choice tests, a skin odor source had the highest valence for landing, followed by a combination of heat and a visual cue, and finally heat and visual cues presented separately. We also measured the durations of the landings, though no significant differences were found.
Similar content being viewed by others


Introduction
Female Aedes aegypti mosquitoes vector dengue, Zika, chikungunya, and yellow fever viruses by repetitive feeding on humans. Skin odor is thought to be the cue that female Ae. aegypti and other anthropophilic mosquitoes use to discriminate humans from other endothermic vertebrates (Gouck 1972; Takken et al. 1997; Dekker et al. 2001, 2002; Besansky et al. 2004; McBride 2016). Aedes aegypti, following an encounter with an above-ambient concentration of CO2, use skin odor to pinpoint a landing site suitable for a blood meal.
Due to the special role of skin odor, we presented Ae. aegypti with skin odor without co-located visual cues. We also quantified landing, a behavior necessary for mosquito blood feeding. We found that Ae. aegypti landed on a source of skin odor presented without a co-located visual cue more frequently than on the visual cue, a heat cue, or even a heated visual cue. The primacy of skin odor contrasts with the view that this diurnal mosquito relies primarily on visual cues during host seeking after navigating upwind along a CO2 plume (van Breugel et al. 2015). Unlike prior work with mosquitoes and other insects (Goodman 1960; Srinivasan and Zhang 1997; Srinivasan et al. 2000; van Bruegel and Dickinson 2012; Parker et al. 2015), in which visual cues elicited landing or persistent nearby flight, we presented heat and skin odor without co-located visual cues. Mosquitoes nonetheless landed on both visually indistinct source of skin odor and, less frequently, heat stimuli.
To assess the relative valence of host-seeking cues used by female Ae. aegypti, we presented skin odor, heat and visual cues in a free-flight wind tunnel, which allowed us to separate a CO2 plume from other host cues. Naïve Ae. aegypti were first exposed to an above-ambient concentration of CO2 at the tunnel’s downwind end and then offered a choice of two competing stimuli on the wind-tunnel floor 50 cm upwind of the release cage. This choice allowed us to determine which cues host-seeking Ae. aegypti preferred. We also tallied the durations of the landings but found no significant differences across cue combinations.
Materials and Methods
Insects
We used the “Orlando” strain of Ae. aegypti (Kuno 2010). The colony was maintained in a L:D 14:10 h cycle, at 25 °C and 70% RH in the UCR Insectary and Quarantine Facility. The females used for colony maintenance were fed defibrinated bovine blood through an artificial membrane (HemoStat Laboratories, Dixon, CA, USA). Larvae were reared in plastic containers and fed TetraMin Tropical Tablets (Tetra Holding GmbH, Melle, Germany). Approximately 50 larvae were reared in each container. All pupae (male and female) from three containers were allowed to emerge into screen cages (BugDorm 30 × 30 × 30 cm, MegaView Science Co., Ltd., Taichung, Taiwan) containing 10% (v) sucrose solution provided ad libitum. All mosquitoes were assumed to have mated and were used only once. Five female mosquitoes, 3–9 days post eclosion, were transferred to cylindrical acrylic release cages (7 × 8 cm i.d.) three hours before the start of assays. Aedes aegypti were assayed 4–8 h into their photophase.
Wind Tunnel
The flight and landing of mosquitoes were observed in a glass wind tunnel 122 × 30.5 × 30.5 cm (Fig. 1A). The exterior of the wind-tunnel floor was covered with black construction paper. Yellow tape was applied to the outside of the glass sidewalls in “x” patterns to provide optomotor feedback (shown only on one side in Fig. 1A). Additional visual feedback was available from the wind tunnel’s structural components and the room external to the wind tunnel. The mosquitoes were therefore not in a featureless visual surround. Air was drawn into the wind tunnel from an adjacent uninhabited room (25 °C and 70% RH). The experimenter did not breathe while loading the release cage into the wind tunnel each trial, so that mosquitoes being transferred into the tunnel were not exposed to a human exhalation of CO2. Airspeed throughout the wind tunnel was 0.2 m/s. The mosquitoes were recorded for 6 min using a video camera (ICD 48, 6 mm lens; Ikegami, Maywood, NJ, USA) positioned 50 cm above the wind tunnel. This allowed observation of most of the tunnel, including the entire area in which cues were presented. Illumination was provided by four infrared LED light banks (AXIS T90A, 850 nm, Axis Communications AB, Lund, Sweden) mounted behind a stainless-steel screen at the downwind end of the wind tunnel. Diffuse room light, provided by incandescent bulbs, measured at ~ 14 lux in the tunnel. The visible spectrum lights were aimed at the junction of the wall and ceiling opposite the wind tunnel. Luminance was measured from a point centered in the wind tunnel and 70 cm from its upwind end, with a Gossen Ultra-Pro (GOSSEN GmbH, Nuremberg, Germany). The luminance at the downwind end was 4 cd/m2, the upwind end 1 cd/m2, the room 4 cd/m2, the wall 8 cd/m2, and the beads 0.067 cd/m2.

(A) Diagram of wind tunnel. Wind flow is from right to left. (B&C) The x-axes go right to left, to match the physical arrangement of the wind tunnel. (B) Carbon dioxide concentration in parts per million (ppm) centered in the tunnel at a height of 14 cm (purple circles) and 1 cm (green squares). The dashed horizontal blue line represents concurrent measurements of the ambient CO2 concentration. (C) The air temperature downwind of the heat pad at three heights. A height of 1 cm is shown in red (squares), 2 cm in orange (circles), and 3 cm in blue (triangles). Concurrent measurements of the ambient air temperature are shown in purple
A visually indistinguishable layer of beads allowed heat or skin odor to be presented independent of a co-located visual cue. The two cue presentation areas in each trial were arranged in the middle of the aluminum pan 13.5 cm from the upwind and downwind ends of the aluminum pan and with 5 cm separating each cue from each other and from the lateral edges of the pan (Fig. 1A). This consistency of location was crucial because the beads were not visibly distinguishable. The pan was placed with its downwind edge 70 cm from the downwind.
To simulate the presence of an upwind vertebrate host, 100 ml/minute of CO2 at 4% concentration mixed with tank air was carried to the wind tunnel via a 3-m-long Tygon® tube, ensuring temperature equilibration. The tube was connected to a glass, L-shaped tube (OD 5.5 mm, ID 3.5 mm) that descended 15 cm from the ceiling on the wind tunnel and extended 20 cm downwind to 60 cm upwind from the release cage. The 4% CO2 mix exited the inner opening at ~ 0.4 m/s, but there was no detectable difference in airspeed 1 cm downwind of the CO2 release point (Omega HHF 52 anemometer, Omega Engineering, Inc., Stamford, CT, USA) and the temperature was identical to the air in the wind tunnel (to within 0.1 °C, same device). The CO2 release tube was centered so that the generated plume of CO2 would engulf the release cage. The CO2 plume was turbulent enough to produce the distinct packets of CO2 needed to elicit upwind flight (Dekker and Cardé 2011) yet compact enough not to mingle with the skin odor plume below. The CO2 plume structure was verified with a visible “smoke” plume of titanium dioxide and hydrochloric acid produced by the reaction of TiCl4 with damp air.
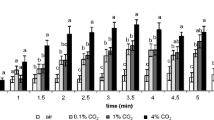
We measured the CO2 concentration with a GasHound CO2 detector (Model LI-800, LI-Core, Nebraska, USA) at 10 points in line with the CO2 source as well as 10 points along the floor (Fig. 1B). Carbon dioxide concentrations were recorded after one minute of equilibration. However, because the gas being sampled is drawn through a tube, a pump, and a filter, this instrument produces time-averaged values in contrast to the nearly instantaneous sensing of CO2 by mosquitoes (Dekker and Cardé 2011). The dashed blue line in Fig. 1B shows that the CO2 did not extend to within 1 cm of the floor 25 cm downwind of the other cue presentation areas. The heights of CO2 measurements refer to height above the beads. The height was kept the same for measurements downwind of the beads. For measurements downwind of the beads this means the true height was 0.7 cm above the tunnel floor. To land on a floor cue the mosquito must fly both vertically down and upwind of the CO2 source. A few mosquitoes approached the glass CO2 release tube. Some cast a few centimeters downwind for several seconds, and others briefly landed on the tip of the CO2 release tube, but they were not scored.
The fluctuating concentration of CO2 at the release cage was expected to elicit take off and to sensitize the mosquitoes to other cues. Because CO2 flux occurs in the field, we did not conduct a CO2 free control. If we had eliminated CO2, the lower take off rate would have made the sample size small for a reason that is not biologically meaningful. The separation of the CO2 plume vertically from the skin odor and heat plumes showed that the cues do not need to be encountered simultaneously.
To ensure that elevated concentrations of CO2 were not present in other parts of the wind tunnel, we took additional readings with a portable Amprobe CO2-100 m (Amprobe, Everett, WA, USA). The readings are shown graphically in Fig. 2.

Plot of the concentration of carbon dioxide at several locations in the wind tunnel under assay conditions. The color of each dot varies from light yellow (406 ppm) to dark purple (4860 ppm), with intermediate concentrations shown in shades of yellow mixed with purple. The lack of dark purple dots anywhere other than directly downwind of the CO2 release point shows the separation of the cues available to the mosquito at any point in its flight. The x-axis shows the distance downwind from the upwind end of the tunnel in centimeters. The y- and z-axes show the displacement from the floor and room-side tunnel wall in centimeters
Cues Presented on the Floor
The orientation of all cues presented on the floor was assigned randomly at the start of each day. To account for possible position effects, their position was switched halfway through the assays being conducted of that type on that day. The beads were put in the same place each time.
Odor Cue
Human skin odor was collected onto black glass beads (12/0 Czech Glass Seed, approximately 2 mm OD toroidal, Precosia Ornela, Zásada, Czech Republic)) by placing 25 ml of beads into a polyester/cotton blend sock, which was then worn by a volunteer (two males and one female) for three hours (Bernier et al. 1999). One volunteer was used per day. Volunteers refrained from alcohol, spicy foods, vigorous exercise, and scented products for three days before and while wearing the beads. The unavoidable variation in human odor on a day-to-day basis was accounted for by statistical blocking.
The beads treated with skin odor were poured into a 55-mm-diameter plastic Petri dish. Beads were allowed to air dry for one hour before assays. The bead-filled Petri dish was then covered with a black aluminum pan (Fig. 3A). While pressing the Petri dish into the pan, the whole arrangement was flipped (Fig. 3B). Clean beads, otherwise identical to those used to collect odor, were then poured into the aluminum pan (Fig. 3C). Removal of the Petri dish produced a visually indistinguishable patch of beads treated with skin odor surrounded by clean beads (Fig. 3D). The area of clean beads within which landing was scored was equal in size (55 mm diameter) to the odoriferous beads. It was crosswind and opposite to the odoriferous beads.

Odor cues were presented on a 5-mm layer of black glass beads (12/0 Czech Glass Seed, approximately 2 mm OD toroidal, Precosia Ornela, Zásada, Czech Republic) in a 32.5 by 26.5 cm black aluminum pan. (A) The empty aluminum pan. (B) The aluminum pan with a Petri dish containing odor beads. (C) The aluminum pan with inverted Petri dish containing odor beads surrounded by clean beads without odor. (D) Petri dish removed leaving no visual target of odor beads within the clean beads
Between use, beads were sonicated with detergent (Micro-90; Aldrich; St. Louis MO, USA), rinsed with distilled water, rinsed with acetone, and baked at 200 °C for 8 h. One week before assays were conducted, the aluminum pans used to hold the beads (32.5 × 26.5 × 1.6 cm, Catering Tray Lid, Smart & Final, Commerce, CA, USA) were spray painted flat black (2X Ultracover, Rust-Oleum Corp. Vernon Hills, IL, USA) and baked at 150 °C for 48 h. Pans containing beads with odor were used for one series of assays with the same treatment combination and then discarded.
A heated odor stimulus was not provided, because heat would change the release rates of the skin odor compounds. The odor released by a heated skin odor cue would not have been comparable to unheated odor.
Heat Cue
A 74-mm diameter silicone heating pad (Cole-Parmer, Vernon Hills, IL, USA) was placed in a black aluminum pan and covered with glass beads to a depth of 5 mm. The surface temperature of the beads measured with a thermocouple (BAT-12, Sensorteck Inc., Clifton, NJ) was 34 °C. The power cord was run through an incision in the bottom of the aluminum pan. The heating pad was not visible from above.
Visual Cue
To add a visual cue, we cut a white tissue (Kimwipe®) into an annulus with a 75 mm outer diameter and a 55 mm inner diameter and placed it atop the beads. The resulting 55-mm-inner black circle was similar in size to visual cues used in some mosquito traps (Bidlingmayer 1994) and wind-tunnel studies (van Breugel et al. 2015). To address concerns that the white Kimwipe® was too reflective, the light gray annuli were cut out of construction paper (Staples Pastel Gray, Staples Inc. Framingham, MA, USA) that was baked for 4 days at 200 °C. To create a facsimile of the visual cue used by van Breugel et al. (2015) an IR-filter (Wratten 2 No. 87, Eastman Kodak Company, Rochester, NY, USA) circle was placed atop a rectangle of gray construction paper that covered half of the bead-filled pan. The inner circles of both annular visual cues were 55 mm in diameter. This matched the diameter of our skin odor cue.
Visual Cue Appearance to the Mosquitoes
Aedes aegypti eyes have a minimum resolvable angle of 12.3 °, allowing them to discern an object approximately the size of human-height from ~ 7 m away (Muir et al. 1992). Depending on the angle of approach, the black inner circle of our white annulus should be discernible up to 26 cm away. However, because the inner and outer edges of the annulus present ellipses that are narrower from certain angles of approach, the maximum discernible distance varies based on angle of approach. The annulus may be detectable to the mosquito beyond these ranges but would be visualized as shades of gray on several ommatidia. Despite the presumed detectability of the visual cue, we observed few landings on unheated visual cues.
The visual cues obscured mosquitoes passing over them, which prevented the use of computer vision. Additionally, when a mosquito flew over the annulus and was not visible on the inner beads nor the other side, it was assumed to have landed on the paper and was scored.
Assay-Pairing Strategy
We conducted two sets of assays daily. The first was a two-choice assay in which skin odor was presented alongside another cue (i.e., heat, visual, or heated-visual cue). The second assay, a single-choice assay, presented the same non-odor cue without a competing odor stimulus. This latter assay served as a reference for determining the extent to which mosquitoes are attracted to visually indistinct heat, visual, and heated visual cues. The order of the assays was alternated daily. Mosquitoes used on a given day were from the same emergence cohort. Each treatment combination was tested across a set of several days.
Cue Presentation
Treatments were presented on a 5-mm layer of black glass beads in a 32.5 by 26.5 cm black aluminum pan. The visually homogeneous layer of beads allowed heat or skin odor to be presented independent of a co-located visual cue (Fig. 3D).
In all trials, a 100 ml/min plume of 4% CO2 and 96% tank air was introduced at the same height as the mosquito release cage to simulate the presence of an upwind vertebrate host. This plume was separated from all cues presented on the tunnel floor (Figs. 1B and 2).
Bioassay Procedure
Five female Ae. aegypti were transferred to release cages 3 h before assays. The mosquitoes were allowed one minute to acclimate to the wind tunnel. The release cage was opened at the upwind end and the mosquitoes were allowed to fly freely under video observation for 6 min.
Data Collection
Landings were scored when a mosquito stopped movement on one of the 55-mm-diameter cues. Individual mosquitoes are visually indistinguishable and potentially could land multiple times on one or both treatments. All landings on the cues during the 6-minute observation period were scored manually with BORIS v.5.1.0 (Friard and Gamba 2016). All data manipulation and statistical tests were conducted using R v.3.5.0 (R Core Team 2013; RStudio Team 2015). The times of the following events were recorded: opening of the release cage, and latencies of takeoff, landing upon, and departure from either of the two cue presentation areas. This allowed for the calculation of the duration of each landing. Flying mosquitoes were indistinguishable on an individual basis. Therefore, “take off” considers only the first mosquito to take off.
Statistical Analysis
The counts on each treatment type were summed by trial and entered into a Wilcoxon signed-rank test. This test treated each trial as a block. Because only one odor source volunteer was used each day, this was de facto blocking by odor source. This statistical blocking also accounts for daily variations in volunteer odor.
The Wilcoxon signed-rank test has an assumption that mosquito choices are independent events. This means that an animal choosing one option cannot subsequently choose another. Our free-flight wind tunnel allows a mosquito to land on one cue presentation area, take off, and land again on the other cue presentation area. The few observed occurrences of landing on both cues were noted and not used in our analyses.
As we cannot keep track of individual mosquitoes and the camera field of view does not fully cover the assay chamber, a few mosquitoes may have taken off from one cue presentation area, flown out of field of view or into glare, and then landed on the other area. Because only a handful of mosquitoes landed on both cue areas within the camera field of view, we believe that only a small percentage of mosquitoes left the field of view and returned to a different cue presentation area. Therefore, to meet the independence assumptions of the Wilcoxon signed-rank test, 5% of landings were trimmed from trials in which both cue presentation areas elicited at least one mosquito landing. A 5% trim was chosen because we believe the percentage of mosquitoes that completed two landings on different cues without being observed and discounted was less than 5%. This trimming was conducted out of an abundance of caution and the trim level was selected a priori. To be invalid, more than one in twenty landings must have been by a mosquito that conducted an improbable maneuver. It is analogous to the “trimmed mean” method described by Tukey and McLaughlin (1963) which is still in common usage (Zhou et al. 2014). Conducting assays of single mosquitoes would have eliminated this problem but would have quintupled the number of assays required from 520 to 2600. We have included Wilcoxon signed-rank test outputs generated without the use of this trimming. None of the probability values change across the 0.05 threshold of significance (Table S1).
Results
We show that a small source of human skin odor in a larger homogeneous visual background elicited far more mosquito landings than a similarly sized heat (p < 0.001), visual (p < 0.001), or visually distinct heat cue (p < 0.001). The CO2 plume was vertically separated from the visual, heat and skin-odor cues, so that at the moment of landing, the choice of stimulus was independent of concurrent sensing of fluctuations in CO2 concentration.
Our study shows that Ae. aegypti, following an encounter with an above-ambient concentration of CO2 as occurs upon approach to a human, uses skin odor to pinpoint a landing site suitable for a blood meal. We found that this diurnal mosquito chooses skin odor over heat, and heat over visual cues.
The y-axes of the plots are adjusted by the number of mosquitoes flown. This makes the bar graphs visually comparable within lettered sections. This concession for accuracy does make it harder to view the low numbers of landings on some treatments. A table of the exact data is available (https://github.com/bendemasisumner/Primacy-of-Human-Odors-Aedes-aegypti).
Mosquitoes landed on skin odor far more frequently than any other stimulus. The heated visual cue and visually indistinct heat cue elicited intermediate numbers of landings. Although the landings are not statistically comparable across groups, the unheated visual cue elicited the fewest landings (Fig. 4C).

Landing counts within a lettered box were by mosquitoes from the same rearing cohorts and were flown on the same set of days. Y-axes are adjusted, within each set of days, in proportion to number assayed. This means the bar graphs are comparable within each lettered section. The calculated probabilities (p-values) above the bars show differences within trial type, not between types of trials (Wilcoxon signed-rank test)
Of 450 mosquitoes given a choice of skin odor or a heat cue, there were 369 landings on skin odor and 115 landings on the heat cue (p < 0.001). On the same days, 385 mosquitoes were assayed with unheated clean beads versus a heat cue; there were 4 landings on the unheated clean beads and 54 landings on the heated beads (p < 0.001) (Fig. 4A).
Of 195 mosquitoes assayed with skin odor versus a heated visual cue, there were 66 landings on skin odor and 10 landings on the heated visual cue (p < 0.001). On the same days, 270 were assayed with unheated clean beads and a heated visual cue. There were only 3 landings on the unheated clean beads while there were 24 landings on the heated visual cue (p = 0.023) (Fig. 4B).
We assayed 440 mosquitoes with skin odor versus a visual cue. The skin odor elicited 427 landings while there were 10 landings on the visual cue (p < 0.001). On those same days, 235 mosquitoes were assayed with unheated clean beads and a visual cue. There was only one landing on the unheated clean beads and 6 landings on the visual cue [not significantly different (p = 0.203) (Fig. 4C)].
We assayed 415 individuals with a heat cue and a visual cue. The heat cue elicited 88 landings while the visual cue elicited only 3 landings (p < 0.001). On those same days, 165 individuals were presented with a heat cue which elicited 20 landings and a heated visual cue which elicited 28 landings [not significantly different (p = 0.222) (Fig. 4D)].
A light gray annulus alone elicited fewer landings than a visually indistinct heat cue (p = 0.003), and a heated light gray annulus did not elicit any more landings than a heated visually indistinct heat cue (p = 0.729) (Fig. 4E).
To verify that our visual cue was not “repellent,” we compared a heated white annulus to a heated light gray annulus. There was no significant difference in landing (Fig. 4F). We found no difference in the number of landings elicited by our light gray annulus against our copy of the IR-filter used by by van Bruegel et al. (2015) (Fig. 4G). We tested our black glass bead visual cue against an IR-filter as used by van Bruegel et al. (2015). Again, there was no difference in landing (Fig. 4H). In this case we provided a neutral gray background that covered one side of the pan. None of unheated visual cues elicited significantly more landings than clean beads.
To address the possibility that differences between our visual cue and those those used by other researchers were leading to a false negative, we replicated the black disk used by van Bruegel et al. (2015), and presented it, 5 mosquitoes at a time, to 390 mosquitoes. Only two landed on it, although many flew near the disk. We also tested a black circle of beads exposed through a hole cut into gray paper. This provided a small black cue in a larger area of neutral color yet still did not elicit landings (Figs. 4H and 5H). This means that all of or visual cues provided on the floor, which were approximately similar to those tested in the literature, failed to elicit substantial mosquito landing.

Landing durations within a letter were by mosquitoes flown on the same days. The p-values in the dot plots show differences within trial type, not between the two types of trials in each lettered section (Kruskal-Wallis test, no correction). The x-axis in each graph is the treatment presented. The lateral displacement of the dots is to ensure that the dot does not overlap with its neighbors. The more landings had a given duration, the more lateral displacement was graphically required. The red triangles indicate the median. A p-value of “NA” indicates an insufficient sample size
Landing Durations
Most pairs of cues, even those in which one elicited vastly more landings than the other, did not have significantly different landing durations (Fig. 5A, B, and C). Some of these were not statistically comparable due to insufficient sample size for at least one cue (Fig. 5C, G, and H). There were no statistical differences between heat versus heated visual cues (Fig. 5D and G). This was mirrored by the lack of significant difference between durations of landings on heat versus light gray annulus (Fig. 5E).
Discussion
Host-seeking in mosquitoes is traditionally understood to be a series of sequential orientation maneuvers. As a mosquito approaches a host, it encounters cues produced by the host, that release a series of discrete maneuvers (Cardé and Willis 2008; Cardé and Gibson 2010; Lacey and Cardé 2011; van Breugel et al. 2015; Cardé 2015). The maneuvers include flight upwind along plumes laden with host cues and orientation toward visual cues. In this sequential-distance model, the mosquito is presumed to encounter progressively more host-specific cues, on the assumption that additional cues that are host-specific become detectable as the mosquito closes the distance to the host. The sequential-distance model does not explain choice when more than one cue is available. Aedes aegypti land on a heat source when CO2 is elevated above background, but rarely on the CO2 source itself (Lacey et al. 2014; McMenamin et al. 2014). The malaria mosquito Anopheles coluzzii landed on a nylon mesh patch imbued with skin odor rather than a CO2 source provided several centimeters away. However, the mosquitoes landed on the mesh only when a plume of elevated CO2 concentration was present (Webster et al. 2015). In both species (Lacey et al. 2014; McMenamin et al. 2014; Webster et al. 2015), the mosquitoes choose to land on the skin odor or heat over the CO2 source.
Other versions of the sequential-distance model suggest that the pairing of cues in addition to the distance between a mosquito and its host also plays a part in determining the mosquito’s response. For instance, one cue may lower the response threshold for another such as skin odor or visual cues (Dekker et al. 2005; van Breugel et al. 2015; Cardé 2015). We propose that in addition to host seeking based on pairs of cues that are detectable a given point, Ae. aegypti has an innate hierarchy of cue preference.
Mosquitoes track odor plumes upwind toward their source by optomotor aenemotaxis (Kennedy 1940; Dekker and Cardé 2011). How far mosquitoes are able to track odor plumes is not precisely known. Using rings of traps around a source of ~ 1 L/min CO2, which is equivalent to ~ 4 sedentary adult humans (Snow 1970; Schreck et al. 1972) captured mosquitoes from many genera, including Aedes. Of these, 92% were captured within 18 m of the CO2 source, suggesting that a human CO2 plume would be attractive to mosquitoes within less than ~ 18 m. This finding has a source of bias, as distant traps cover a smaller angle from the CO2 source. This may have provided an underestimate of the distance of attraction.
After following a plume of CO2, the sequential-distance model assumes that host-seeking Aedes mosquitoes orient toward visual cues (Cardé and Gibson 2010; van Breugel et al. 2015). The eyes of Ae. aegypti have a minimum resolvable angle of 12.3 °, allowing them to discern a human-sized object from ~ 7 m away (Muir et al. 1992). The black inner circle of our white annulus would be discernible up to 26 cm away, depending on the angle of approach. However, we observed very few landings on unheated visual cues (Fig. 4C, D, E, G, and H). Although the visual cue provided was obviously different from that provided by a human, it was similar to those presented by other researchers (van Breugel et al. 2015).
The scarcity of landings on our unheated visual cues contrasts with the finding of van Bruegel et al. (2015) that Ae. aegypti spent more time flying near a visual than a heat cue. However, they did not report whether the mosquitoes that flew within “an 8 × 8 × 4 cm volume above and downwind” of the black disk, made out of near-infrared transparent plastic, actually landed on it. Others have observed that An. coluzzii often fly near visual cues without landing on them (Hawkes and Gibson 2016).
Liu and Vosshall (2019) found that magnetically tethered Ae. aegypti orient toward black vertical stripes, which would support that visual cues are important prior to landing. They also quantified mosquito occupancy, i.e., landings and remaining landed, after landing on an unheated visual cue. Although a vertically oriented black dot elicited more mosquito occupancy than the surrounding white paper, it was less than half of mosquito occupancy of heat without a visual cue.
Our results affirm that even in the presence of elevated CO2, visual cues elicit few landings. Few mosquitoes landed on the small, clean, unheated visual cues. A close mimic of human visual cues would have been preferred but would have interfered with the presentation of other cues. The minimal response to the alternative visual cues is shown in Fig. 4H and 5H. Because landing is a prerequisite for blood feeding and therefore pathogen transmission, it is an important diagnostic measure of mosquito host-seeking behavior.
In studies of insect landing in which stereotyped maneuvers precede coming into contact with the substrate, insects were provided with a distinct visual cue. Leg extension, body turns (saccades) and pitch change (van Breugel and Dickinson 2012) are triggered by objects covering an expanding portion of the insect visual field. Honey bees can maintain a fixed angle relative to a visual cue in order to execute a smooth descent path toward and landing upon the cue (Srinivasan and Zhang 1997; Srinivasan et al. 2000). In contrast to those studies with prominent visual cues, we presented heat and skin odor without co-located visual cues. The visual cues available to the mosquito for optomotor feedback were lateral and above the mosquito (Fig. 2A). Our mosquitoes still executed landings, using a mechanism different from those previously studied.
In addition to following odor plumes, Ae. aegypti at close range can follow convective heat plumes. Aedes aegypti landed frequently on a nylon mesh cone placed 5 cm above a 43.3 °C black billiard ball but failed to land when the convection was interrupted by a long-wave, infrared-transparent KRS-5 filter (Peterson and Brown 1951). The tips of the antennae of Ae. aegypti are equipped with neurons housed in coeloconic sensilla that exhibit phasic shifts in response to air temperature changes as small as 0.05 °C (Davis and Sokolove 1975). The human-temperature heating pad in our assay produced a plume 0.2 °C above ambient ~ 30 cm downwind. Therefore, the heat plume could have been detectable to the mosquitoes throughout a large swath of the wind tunnel and yet they still preferred to land on a source of skin odor.
The distance at which skin odor plumes are detectable to a mosquito in the field is unclear. Dekker and Cardé (2011) found that skin odor, supplied by an odor stream from an enclosed arm ~ 1 m upwind, readily elicited upwind flight of Ae. aegypti. However, when the skin odor plume was diluted five-fold, it elicited proportionally fewer flights. When a plume of CO2 was added, the mosquitoes were “instantly sensitized” to the diluted skin odor and surged upwind (Dekker and Cardé 2011). Such sensitization clearly could have occurred in our trials, and also would have contributed to landing on a skin odor patch in An. coluzzii (Webster et al. 2015).
Skin odor is thought to be the cue that anthropophilic mosquitoes use to tell humans apart from other endothermic vertebrate hosts (Gouck 1972; Takken et al. 1997; Pates et al. 2001; Dekker et al. 2001, 2002; Besansky et al. 2004; McBride 2016; DeGennaro et al. 2013) found that orco mutant Ae. aegypti lose their strong preference for human odor over that of guinea pig in a cage assay. By lacking orco, the olfactory coreceptor, these mosquitoes lost the function of all of their olfactory receptors. Anopheline mosquitoes (Ribbands 1946) and Ae. aegypti (Trpis and Hausermann 1978) also appear to use human scent for house-entering, which can occur well before biting. Once inside a human dwelling, host seeking could be triggered by a fluctuating concentration of CO2 (Dekker and Cardé 2011). Our findings show that human skin odor not only provides a human-specific cue, but also provides a direct host-seeking and landing cue.
Landing durations were contrasted with a Kruskal-Wallis test. Because the number of landings on a cue is, by definition, the sample size of landing duration, the differences in landing counts on different cues may have limited the statistical power of our test and may explain some of the lack of significant differences between landing durations. This is not unexpected, because the experiment was designed to show a difference in the number of landings on different cues. Landing and remaining landed are separate behavioral categories. Landing duration is a potential measure of cue salience. Mosquitoes have chemoreceptors on their tarsi (Sparks et al. 2013) and labella (Saveer et al. 2018). Once a mosquito’s tarsi or labella contact beads treated with skin odor, the mosquito might detect non-volatile chemicals such as amino acids.
Conclusions
Here we establish that following exposure to a plume of CO2, heat stimulus, and a heated visual cue, all evoke at least some orientation and landing. However, the use of a choice bioassay allowed these cues to be ranked in a valance hierarchy. Naïve Ae. aegypti had a high preference for landing on a skin odor source; heated cues were the second most effective cue for eliciting landing, whereas the unheated visual cues elicited virtually no landings. This demonstrates that Ae. aegypti, a day-biting mosquito, are able to locate and land on skin odor and heat without a co-located visual cue. The order, primacy, and interaction of cues used by Ae. aegypti and other mosquitoes during host finding in the field remain to be firmly established. However, our findings suggest that Ae. aegypti track and land on cues based on an innate hierarchy. This hierarchy appears to rank cue types in descending order of human specificity. The primacy of skin odor contrasts with the view that this diurnal mosquito relies primarily on vision to find and potentially land on a host following detection of a fluctuating concentration of CO2.
Data Availability
GitHub link to follow.
Code Availability
GitHub link to follow.
References
Bernier UR, Booth MM, Yost RA (1999) Analysis of human skin emanations by gas chromatography/mass spectrometry. 1. Thermal desorption of attractants for the yellow fever mosquito (Aedes aegypti) from handled glass beads. Anal Chem 71:1–7
Besansky N, Hill C, Costantini C (2004) No accounting for taste: host preference in malaria vectors. Trends Parasitol 20:249–251
Bidlingmayer WL (1994) How mosquitoes see traps: role of visual responses. J Am Mosq Control Assoc 10:272–279
Cardé RT (2015) Multi-cue integration: how female mosquitoes locate a human host. Curr Biol 18:21–25
Cardé RT, Willis MA (2008) Navigational strategies used by insects to find distant, wind-borne sources of odor. J Chem Ecol 34:854–866
Cardé RT, Gibson G (2010) Host finding by female mosquitoes: mechanisms of orientation to host odours and other cues. In: Takken W, Knols BGJ (eds) Olfaction in Vector-Host Interactions. Wageningen Academic Publishers, Wageningen, pp 115–140
Davis E, Sokolove P (1975) Temperature responses of antennal receptors of the mosquito, Aedes aegypti J Comp Physiol 96:223–236
DeGennaro M, McBride CS, Seeholzer L, Nakagawa T, Dennis EJ, Goldman C, Jasinskiene N, James AA, Vosshall LB (2013) orco mutant mosquitoes lose strong preference for humans and are not repelled by volatile DEET. Nature 7455:487–491
Dekker T, Cardé RT (2011) Moment-to-moment flight manoeuvres of the female yellow fever mosquito (Aedes aegypti L.) in response to plumes of carbon dioxide and human skin odour. J Exp Biol 214:3480–3494
Dekker T, Takken W, Cardé RT (2001) Structure of host-odour plumes influences catch of Anopheles gambiae s.s. and Aedes aegypti in a dual-choice olfactometer. Physiol Entomol 26:124–134
Dekker T, Steib B, Cardé RT, Geier M (2002) L-lactic acid: a human-signifying host cue for the anthropophilic mosquito Anopheles gambiae Med Vet Entomol 16:91–98
Dekker T, Geier M, Cardé RT (2005) Carbon dioxide instantly sensitizes female yellow fever mosquitoes to human skin odours. J Exp Biol 208:2963–2972
Friard O, Gamba M (2016) BORIS: a free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol Evol 7:1324–1330
Goodman LJ (1960) The landing responses of insects I. The landing response of the fly Lucilia sericata, and other Calliphorinae. J Exp Biol 37:854–878
Gouck HK (1972) Host preferences of various strains of Aedes aegypti and Aedes simpsoni as determined by an olfactometer. B World Health Organ 47:680–683
Hawkes F, Gibson G (2016) Seeing is believing: the nocturnal malarial mosquito Anopheles coluzzii responds to visual host-cues when odour indicates a host is nearby. Parasite Vector 9:320
Kennedy JS (1940) The visual responses of flying mosquitoes. Proc Zool Soc Lond Ser A 109:221–242
Kuno G (2010) Early history of laboratory breeding of Aedes aegypti (Diptera: Culicidae) focusing on the origins and use of selected strains. J Med Entomol 47:957–971
Lacey ES, Cardé RT (2011) Activation, orientation and landing of female Culex quinquefasciatus in response to carbon dioxide and odour from human feet: 3D flight analysis in a wind tunnel. Med Vet Entomol 25:94–103
Lacey ES, Ray A, Cardé RT (2014) Close encounters: contributions of carbon dioxide and human skin odour to finding and landing on a host in Aedes aegypti Physiol Entomol 39:60–68
Liu MZ, Vosshall LB (2019) General visual and contingent thermal cues interact to elicit attraction in Female Aedes aegypti mosquitoes. Curr Biol 29:2250–2257
McBride CS (2016) Genes and odors underlying the recent evolution of mosquito preference for humans. Curr Biol 26:41–46
McMenamin CJ, Corfas RA, Matthews BJ, Ritchie SA, Vosshall LB (2014) Multimodal integration of carbon dioxide and other sensory cues drives mosquito attraction to humans. Cell 156:1060–1071
Muir LE, Thorne MJ, Kay BH (1992) Aedes aegypti (Diptera: Culicidae) vision: spectral sensitivity and other perceptual parameters of the female eye. J Med Entomol 29:278–281
Parker JE, Angarita-Jaimes N, Abe M, Towers CE, Towers D, McCall PJ (2015) Infrared video tracking of Anopheles gambiae at insecticide-treated bed nets reveals rapid decisive impact after brief localised net contact. Sci Rep-UK 5:13392
Pates HV, Takken W, Stuke K, Curtis CF (2001) Differential behaviour of Anopheles gambiae sensu stricto (Diptera: Culicidae) to human and cow odours in the laboratory. Bull Entomol Res 91:289–296
Peterson DG, Brown AWA (1951) Studies of the responses of the female Aëdes mosquito. Part III. The response of Aëdes aegypti (L.) to a warm body and its radiation. Bull Entomol Res 42:535–541
R Core Team (2013) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. http://www.R-project.org/
Ribbands CR (1946) Moonlight and house-haunting habits of female anophelines in West Africa. Bull Entomol Res 36:395–417
RStudio Team (2015) RStudio: Integrated development for R. RStudio Inc., Boston
Saveer AM, Pitts RJ, Ferguson ST, Zwiebel LJ (2018) Characterization of chemosensory responses on the labellum of the malaria vector mosquito, Anopheles coluzzii Sci Rep 8:5656
Schreck CE, Gouck HK, Posey KH (1972) The range of effectiveness and trapping efficiency of a plexiglas mosquito trap baited with carbon dioxide. Mosq News 32:496–501
Snow WF (1970) The effect of a reduction in expired carbon dioxide on the attractiveness of human subjects to mosquitoes. Bull Entomol Res 60:43–48
Sparks JT, Vinyard BT, Dickens JC (2013) Gustatory receptor expression in the labella and tarsi of Aedes aegypti Insect Biochem Mol Biol 43:1161–1171
Srinivasan MV, Zhang SW (1997) Visual control of honeybee flight. In: Lehrer M (ed) Orientation and Communication in Arthropods. Birkhauser Verlag, Basel, pp 94–113
Srinivasan MV, Zhang SW, Chahl JS, Barth E, Venkatesh S (2000) How honeybees make grazing landings on flat surfaces. Biol Cybern 83:171–183
Takken W, Knols BGJ, Otten H (1997) Interactions between physical and olfactory cues in the host-seeking behaviour of mosquitoes: the role of relative humidity. Ann Trop Med Parasitol 91:S119–S120
Trpis M, Hausermann W (1978) Genetics of house-entering behaviour in East African populations of Aedes aegypti (L.) (Diptera: Culicidae) and its relevance to speciation. Bull Entomol Res 68:521–532
Tukey JW, McLaughlin DH (1963) Less vulnerable confidence and significance procedures for location based on a single sample: trimming/Winsorization. Sankhyā 25:331–352
van Breugel F, Dickinson MH (2012) The visual control of landing and obstacle avoidance in the fruit fly Drosophila melanogaster J Exp Biol 215:1783–1798
van Breugel F, Riffell J, Fairhall A, Dickinson MH (2015) Mosquitoes use vision to associate odor plumes with thermal targets. Curr Biol 25:2123–2129
Webster B, Lacey ES, Cardé RT (2015) Waiting with bated breath: opportunistic orientation to human odor in the malaria mosquito, Anopheles gambiae, is modulated by minute changes in carbon dioxide concentration. J Chem Ecol 41:59–66
Zhou X, Lindsay H, Robinson MD (2014) Robustly detecting differential expression in RNA sequencing data using observation weights. Nucleic Acids Res 42:11
Acknowledgements
We are grateful to Drs. ES Lacey for experimental advice and S Ghosh for statistical guidance. We also thank Drs. JG Millar and AC Gerry for critical reviews of the manuscript.
Funding
We acknowledge funding from the Pacific Southwest Regional Center of Excellence for Vector-Borne Diseases funded by the U.S. Centers for Disease Control and Prevention (Cooperative Agreement 1U01CK000516).
Author information
Authors and Affiliations
Contributions
BDS and RTC designed thee experiments and wrote this paper; BDS carried out these trials and analyzed our findings.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflicts of interest.
Ethics Approval
Not applicable.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 25.6 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sumner, B.D., Cardé, R.T. Primacy of Human Odors Over Visual and Heat Cues in Inducing Landing in Female Aedes aegypti Mosquitoes. J Insect Behav 35, 31–43 (2022). https://doi.org/10.1007/s10905-022-09796-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10905-022-09796-2