
Abstract
Maize (Zea mays L.) is the most produced field crop all over the world. One of its most critical diseases that results in economic loss is ear rot caused by various Fusarium species. Previous researches have shown that polyamines, found in all living cells, play crucial role in biotic stress responses. At the same time, biosynthesis of polyamines is of paramount importance not only for plants but also for their pathogens to promote stress tolerance and pathogenicity. In our study, we investigated the polyamine content changes induced in the seedlings of two maize genotypes of different susceptibility by isolates of Fusarium verticillioides and Fusarium graminearum, two Fusarium species of different lifestyles. Apart from that, it was examined how infection efficiency and changes in polyamine contents were modified by salicylic acid or putrescine seed soaking pre-treatments. Our observations confirmed that initial and stress-induced changes in the polyamine contents are not directly related to tolerance in either coleoptile or radicle. However, the two pathogens with different lifestyles induced remarkably distinct changes in the polyamine contents. The effect of the seed soaking pre-treatments depended on the pathogens and plant resistance as well: both salicylic acid and putrescine seed soaking had positive results against F. verticillioides, while in the case of infection with F. graminearum, seed soaking with distilled water alone affected biomass parameters positively in the tolerant genotype.
Similar content being viewed by others


Introduction
Maize (Zea mays L.) is one of the most important field crops grown all over the world. Regarding the extent of economic losses, its most critical diseases are ear rot and stalk rot, which are caused by toxin-producing Fusarium species (Meissle, 2010). In maize, several different Fusarium species occur and diseases can break out as a result of simultaneous presence of various Fusarium species in the plants (Munkvold 2003; Folcher et al. 2009; Mesterházy et al. 2011). Ear rot (GER) provoked by Fusarium graminearum Schwabe [teleomorph = Gibberella zeae (Schwine)] is a frequent pathogen of rainy years, whereas ear rot (FER) caused by Fusarium verticillioides (F. moniliforme J. Sheld. teleomorph = G. moniliformis Wineland) occurs rather in warm and dry seasons (Vigier et al. 1997; Doohan et al. 2003). The impact of climate change manifests in the growing prevalence of dry and warm weather (Jacob et al. 2014); therefore, FER has an increasing role in Fusarium infections. Fusarium species can infect any parts of the maize plant, from germination stage to harvesting. Pathogens may infect and damage the seeds and seedlings still in the soil, before emergence (Munkvold 2003). Fusarium species can overwinter on plant residues (Nyvall and Kommedahl 1970). Chlamydospores of F. graminearum are viable in the soil for long periods (Sutton 1982); in addition, they even form perithecia, from which they can further infect through ascospore discharge (Guenther and Trial, 2005). F. verticillioides produces thickened hyphae, which also increases its survival in the soil (Maiorani et al. 2008). Both species ensure their pathogenicity by forming a mass of micro- and macroconidia (Munkvold 2003). While F. graminearum rapidly changes to a necrotrophic phase during its life cycle (Quarantin et al. 2019), F. verticillioides is considered a hemibiotroph, as it coexists with maize as an endophyte and can occasionally change to a necrotrophic phase and cause disease at any stages of the plant's life cycle (Beccaccioli et al. 2021).
Polyamines (PA) are aliphatic amines found in all living cells. They play a crucial role in plant development, in the regulation of vital plant functions and in stress responses (Hasanuzzaman et al. 2019; Wang et al. 2019). The most studied and most abundant PAs in plants are putrescine (PUT), spermidine (SPD) and spermine (SPM). Higher PAs, such as SPD and SPM, are synthesized from PUT. During their catabolism, 1, 3 diaminopropane (DAP) forms and hydrogen peroxide releases (Pál et al. 2015). The protective role of PAs during abiotic stresses in various plant species and in maize has already been reported in several studies (Jiménez-Bremont et al. 2007; Gill and Tuteja 2010; Yu et al. 2019; Ramazan et al. 2022). In addition, growing evidence indicates that PAs also play a key role in regulating plant immune response. Accumulation of H2O2 as a result of PA catabolism and of nitric oxide due to induction by SPD/SPM play important signalling roles in plant–pathogen interactions (Hussain et al. 2011). Increased biosynthesis and conversion of PUT to SPD and SPM along with their increased catabolism was evident in the resistant maize lines compared the susceptible line during Aspergillus flavus infection (Majumdar et al. 2019). Transgenic tobacco plants with increased PA oxidase gene expression, exhibited pre-induced resistance towards infections, including biotrophic and hemibiotrophic diseases, too (Moschou et al. 2009). While increased expression level of PA transporter genes has been also reported during F. graminearum infection, as the presence of PAs may be beneficial to both the plant host and fungal pathogen (Harris et al. 2016). Nevertheless, biotrophic and necrotrophic pathogens may induce distinct changes to PAs. Biotrophic pathogens predominantly induce PA synthesis; therefore, the inhibition of PA synthesis and the activation of PA degradation can be important in protecting against them. Necrotrophic infection, however, during the catabolism of the PA accumulated in the apoplast, may be accompanied by hydrogen peroxide release, which enhances the formation of necrotic lesions (Pál and Janda 2017). Although the protective effects of PA treatments under abiotic stress conditions have been confirmed in several cases (Farooq et al. 2008; Cvikrová et al. 2013; Alzahrani and Rady, 2019; Sadeghipour 2019), their impact in pathogenic infections is controversial and poorly studied. For example, SPD and SPM spray treatment in Arabidopsis was effective in promoting long-term resistance against necrotrophic B. cinerea (Janse van Rensburg et al. 2021), whereas in tomato, the increased susceptibility to B. cinerea was reversible by applying PA biosynthesis inhibitors (Handa et al. 2011). There is evidence that effective immune response to plant pathogens is regulated by plant hormones, such as salicylic acid (SA). SA is crucial both in local and systemic defence responses. Systemic acquired resistance (SAR) is an SA-dependent response (Gao et al. 2015). Changes in endogenous SA content and the protective effect of externally administered SA have been reported in several pathogenic infections (Amborabé et al. 2002; Szőke et al. 2011; Qi et al. 2012; Ding et al. 2018; Radojicic et al. 2018; Li et al. 2022), including Fusarium. However, based on the data from literature, there may not only be a connection between the quantitative changes of SA and PAs, but the existence of a positive feedback loop at signal transduction level is becoming more and more evident (Canales et al. 2019; Pál et al. 2021; Szepesi et al. 2022).
Since the role of PA metabolism in the interaction between plants and pathogens with different lifestyles is still not clear, we aimed to study the changes in the PA metabolism induced by the most important ear rot pathogens–the hemibiotrophic F. verticillioides and the necrotrophic F. graminearum –, causing damages to maize worldwide. In addition, we examined whether SA and PUT seed soaking pre-treatments had any positive effect on biomass production against Fusarium infection. In addition, we investigated the effects of the mentioned pre-treatments on the PA levels of plants. In the present research, two maize genotypes with different levels of susceptibility were tested. Measurements were done both in the coleoptile and in the radical as well.
Materials and method
The tested genotypes
The treatments were applied in two different genotypes. Both inbred lines were bred in Martonvásár: EXPMV01 with an Iodent background is susceptible to ear rot, while EXPMV02 is an ear rot-tolerant line with an ISSS background. Seeds were produced in the experimental nursery of the Agricultural Institute, Centre for Agricultural Research (Martonvásár, Hungary, 47◦18' N, 18◦46' E, altitude: 120 m). The crops were harvested in ears at an average moisture content of 23% and were dried in a box dryer to reach 13% moisture content. In the next step, kernels were removed by a maize sheller and were stored at 10 °C, in the warehouse of the institute.
Preparation of pathogen inoculum
For the artificial inoculation, FVTE and IFA-66 isolates of a F. verticillioides (FV) and F. graminearum (FG) were used. Pure cultures were stored in a refrigerator at 4 °C, on a 1:1:1 (v/v/v) mixture of sterilised soil, turf and silica sand. Isolates required for the inoculation were transferred into Petri dishes, onto PDA medium (Leslie and Summerell 2006) and incubated for 5 days at 25 °C, in a Thermo Fisher—Heratherm™ Compact Microbiological Incubator.
Experimental design
The selected genotypes (EXPMV01, EXPMV02) were studied in a two-factor design, in three replicates. One replicate contained 2 × 5 seed samples, which means that one treatment contained 30 samples. Each genotype was characterised with data from a total of 360 seed samples. Petri dishes of 9 cm diameter were poured with 20 ml PDA medium and stored in a laminar box until use. Before treatments, seeds were sterilised using modified instructions of Daniels, (1983) and Szőke et al. (2018): for the disinfection, seeds were soaked for 2 × 5 min in a 50 °C water bath, applying 5-min cooling period between two soaking treatments. Subsequently, seeds were disinfested for 10 min with 10% ethanol solution and after that with 1% Neomagnol solution for 15 min. Finally, the samples were washed twice with distilled water.
After the disinfection, kernels were dried in a laminar box. Batches without soaking (absolute control: ABC) were used after sterilisation. The remaining batches were soaked for 16 h as a pre-treatment with distilled water (DW), 0.5 mM salicylic acid (SA) or 0.5 mM putrescine (PU) solution. Pre-treatment concentrations were determined based on the results of previous experiments (Szalai et al. 2016; Tajti et al. 2018). After drying, batches of 5 kernels were placed in the Petri dishes containing PDA medium, and 5-mm discs containing Fusarium isolates of the relevant type of infection were placed next to the seeds. Altogether, 12 treatments were applied (Table 1). Absolute control treatments: 1. no soaking without infection (ABC); 2. no soaking, infected with F. verticillioides (ABFV); 3. no soaking, infected with F. graminearum (ABFG). Control treatments: 4. soaking with distilled water without infection (DWC); 5. distilled water + F. verticillioides (DWFV); 6. distilled water + F. graminearum (DWFG). Salicylic acid treatments: 7. Soaking with salicylic acid without infection (SAC); 8. salicylic acid + F. verticillioides (SAFV); 9. salicylic acid + F. graminearum (SAFG). Putrescine treatments: 10. Soaking with putrescine without infection (PUC); 11. putrescine + F. verticillioides (PUFV), 12. putrescine + F. graminearum (PUFG). Seeds and fungi were grown for 7 days in laminar box.
Determination of fresh weight
After 7 days, coleoptiles and radicles were collected for each treatment. For each replicate, weights of coleoptiles and radicals were measured using a laboratory scale. Until the quantitative analysis of PAs, the measured samples were stored at −80 °C.
Quantitative analysis of PAs
Extraction and analysis of PAs were carried out using the description of Németh et al. (2002). Using liquid nitrogen and silica sand, 0.2 g samples of coleoptile and radicle were homogenised, then extracted with 2 × 1 ml 0.2 M ice-cold perchloric acid. Afterwards, the samples were left on ice for 20 min and then centrifuged at 10 000 g for 2 min at 4 °C. For the derivatisation, dansyl chloride was used. To 100 μl supernatant, 200 μl saturated sodium carbonate solution and 400 μl dansyl chloride solved in acetone (5 mg ml−1) were added. The reaction mixture was incubated in the dark for 60 min at 60 °C, then 100 μl of proline solution ( 100 mg ml−1) was added and it was further incubated in the dark at room temperature for 30 min. The reaction mix was extracted for 30 s with 500 μl toluene. The top organic layer was pipetted into 1.5 ml Eppendorf tubes using Pasteur pipettes and evaporated to dryness in vacuum. The residue was taken in 1 ml of 100% methanol, then filtered through a PTFE membrane filter with 0.2 μm pore size and analysed with Waters HPLC system (Waters, Milford, MA USA). The HPLC device included an autosampler, a column thermostat and a pump system for gradient mixing, to which a W474 scanning fluorescence detector had been connected. Data were evaluated by Millenium 32 software. 2 μl of the derivatised sample was injected into the Kinetex 5u C18 100A 100 × 2.1 mm column. For gradient separation, two different types of solvents were used [A: 44% acetonitrile (ACN) B: ACN: methanol (MeOH) = 7:3]. During the analysis, the flow rate was 0.5 ml min−1 and the column temperature was 40 °C. PAs derivatised with dansyl chloride were detected with a fluorescence detector at an excitation wavelength of 340 nm and an emission wavelength of 515 nm.
Statistical analysis
For the statistical analysis of the data obtained, Agrobase 99® for Microsoft Windows® (Agronomix Inc.), SPSS 16.0 (IBM Ltd.) and Microsoft Windows® Excel data management software were used. Multi-way ANOVA and post-hoc Duncan test were applied to evaluate the significance of treatments. Results obtained from the statistical probes were interpreted based on the study of Sváb, (1981). Microsoft Windows® Excel data management software was used to arrange the available data for statistical analysis. In the case of Agrobase 99® for Microsoft Windows® statistical software, to organise data, edit diagrams and determine correlation coefficients, built-in modules of MS® Excel data management software were used. ANOVA result tables are presented in Tables 2, 3 and 4.
Results
Pronounced differences in the biomass parameters of the maize genotypes were depended on pathogenic strategies of Fusarium spp.
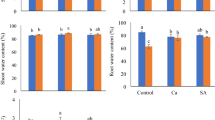
The coleoptile fresh weight (CFW) in the absolute control (ABC) was higher in the susceptible (EXPMV01) genotype, compared to the tolerant one (EXPMV02) (Fig. 1A). The FV treatment (ABFV) slightly increased it, while the FG infection (ABFG) decreased it in both genotypes compared to the control (ABC), which is in accordance with the pathogenic strategies of FV and FG. Seed-priming treatments (DWC, SAC, PUC) alone (i.e., without any Fusarium infection) compared to the control (ABC) did not significantly influence the CFW in line EXPMV01, but they increased it in the tolerant genotype (EXPMV02). Under FV infection the seed soaking treatments further increased the CFW, especially in the susceptible one (EXPMV01), and in the case of SA and PUT treatments compared to the DWC, SAC, PUC or even to ABFV and DWFV treatments. Following the FG infection, the seed pre-treatments (DWFG, SAFG, PUFG) did not affect the susceptible genotype but induced an increase in the tolerant one (EXPMV02) compared to the ABFG treatment alone. However, the greatest increase was found for the DWFG, and lower values were measured for SAFG or PUFG treatments (Fig. 1A) compared to the ABFG. These results showed that the tolerant (EXPMV02) line had more positive responses to the seed pre-treatments, and that these treatments could increase the biomass even under the necrotrophic infection.

Differences in the coleoptile fresh weight A and radicle fresh weight B of the susceptible (EXPMV01) and tolerant (EXPMV02) maize genotypes after different treatments (C: without infection, AB: no soaking, DW: soaking with distilled water, SA: soaking with 0.5 mM salicylic acid, PU: soaking with 0.5 mM putrescine, FV = F. verticillioides infection, FG = F. graminearum infection.) Values are means ± standard error (SE). Different letters indicate statistically significant differences compared to each others (Duncan, p < 0.05)
The radicle fresh weights (RFW) of the two genotypes were similar under absolute control condition (ABC) (Fig. 1B). Although the FV treatment decreased the RFW in both genotypes compared to the control (ABC), it was only statistically significant in the case of the susceptible line (EXPMV01). FG infection also caused significant decrease of the RFW with pronounced lower value for the susceptible (EXPMV01) genotype. Seed soaking treatments (DWC, SAC, PUC) had positive effects on the RFW mainly in the EXPMV02 genotype under either control condition or biotic stress conditions compared to the control (ABC). After FV treatment SAFV and PUFV, after FG infection all the applied pre-treatments (ABFG, DWFG, SAFG, PUFG) had positive effect on the RFW compared to the adequate control (ABFV or ABFG) (Fig. 1B).
Weights of coleoptile and radical were significantly influenced by the genotypes, the treatments, and the interaction between these two factors (p = 0.1%). According to the two-way ANOVA table (Table 2), both coleoptile and radical fresh weights depended mostly on the genotypes. The treatments had 50% less impact on the weight of the coleoptile than the genotype, while in the case of the radicle, the treatments had 20% less influence in comparison to the genotype. The lowest impact was exhibited by the genotypes × treatments (Table 2).
Changes in polyamine contents of the maize seedlings after seed-priming, Fusarium infection and combined treatments
In the absolute control samples (ABC), the levels of PUT and SPM were higher in the tolerant genotypes (EXPMV02) than in the susceptible one (EXPMV01), while the SPD content was slightly higher in the susceptible line both in the coleoptile and radicle (Fig. 2–3.). Comparing the PA content in the coleoptile and radicle, the PUT level was higher in the radicle, but the amounts of SPD, SPM, and DAP were greater in the coleoptile in both genotypes. While in the coleoptile, the SPM was the most abundant PA, in the radicle PUT showed the greatest values in both genotypes (Fig. 2–3.).

Changes in the polyamine contents, namely putrescine (PUT) A, spermidine (SPD) B, spermine (SPM) C and 1,3-diaminopropane (DAP) D, in the coleoptile of the susceptible (EXPMV01) and tolerant (EXPMV02) maize genotypes after different treatments (C: without infection, AB: no soaking, DW: soaking with distilled water, SA: soaking with 0.5 mM salicylic acid, PU: soaking with 0.5 mM putrescine, FV = F. verticillioides infection, FG = F. graminearum infection. Values are means ± standard error (SE). Different letters indicate statistically significantly differences to each others (Duncan, p < 0.05)

Changes in the polyamine contents, namely putrescine (PUT) A, spermidine (SPD) B spermine (SPM) C and 1,3 diaminopropane (DAP) D in the radicles of susceptible (EXPMV01) and tolerant (EXPM,V02) maize genotypes after different treatments (C: without infection, AB: no soaking, DW: soaking with distilled water, SA: soaking with 0.5 mM salicylic acid, PU: soaking with 0.5 mM putrescine, FV = F. verticillioides infection, FG = F. graminearum infection.) Values are means ± standard error (SE). Different letters indicate statistically significant differences to each others (Duncan, p < 0.05)
Markedly distinct impact of F. graminearum infection on the individual polyamine compounds and the effects of seed-priming was found in the coleoptile
The levels of the individual PA compounds were different in the investigated two genotypes already in the ABC samples. The higher DAP level (catabolite product of the higher PAs) in the coleoptile in the tolerant genotype (EXPMV02) suggests a more intensive terminal catabolism, which can be responsible for the overall lower SPD and SPM average levels for the tolerant line (EXPMV02).
Despite the described differences, in most cases, Fusarium infections alone induced similar changes in the PA contents of the two genotypes. Although the ABFV infection did not influence the PUT (Fig. 2A), SPD (Fig. 2B) and DAP (Fig. 2D) levels, it significantly decreased the amount of SPM (Fig. 2C) in both lines compared to the control (ABC). However, the most remarkable changes were found after ABFG infection, namely the PUT and DAP concentrations increased, while the SPD and SPM values decreased compared to the adequate ABC samples in both genotypes. It can be concluded that the effect of ABFG was more pronounced on the PA contents in both genotypes with the highest PUT and DAP, but lowest SPD and SPM levels after infection in the tolerant genotype (EXPMV02).
PUT content (Fig. 2A) was not influenced significantly by the DW, SA or PUT priming without Fusarium infection (DWC, SAC and PUC) in the susceptible genotype, and only SA seed soaking increased it slightly in the tolerant genotype (EXPMV02). When seed-soaking treatments was followed by the FV infection, the PUT level did not change significantly in comparison to the relevant controls (DWC, SAC or PUC) in either genotype. Although the FG infection itself increased the PUT accumulation in both genotypes compared to ABC ones. Among the combined seed soaking and FG treatments, only SAFG treatment could further increase it in the case of the susceptible genotype (EXPMV01) (Fig. 2A).
Under control conditions (ABC, DWC, PUC, SAC), without any Fusarium infection, only the PUT priming could increase slightly, but statistically significantly the SPD content (Fig. 2B) in the susceptible genotype (EXPMV01), while in the tolerant one (EXPMV02) all seed soaking pre-treatments (DWC, SAC and PUC) decreased it compared to the ABC one. During the FV treatment all seed soaking treatments decreased the amount of SPD in both genotypes compared to the ABC or the ABFV-treated samples. Although the FG infection alone decreased the SPD concentrations in both genotypes compared to the absolute controls, they induced different responses in the two lines when the SA and PUT seed soaking were followed by FG treatment. In the susceptible one SA or PUT priming further decreased the SPD level, while in the tolerant one (EXPMV02) the pre-treatment with SA or PUT increased it compared to the ABFG-treated samples (Fig. 2B).
The seed soaking treatments alone induced different changes in the SPM content (Fig. 2C) of the two genotypes. Except for SA pre-treatment, they did not lead to characteristic changes in it in the susceptible genotype (EXPMV01), but they dramatically decreased it in the tolerant one (EXPMV02). Both Fusarium treatments lowered the level of SPM compared to the ABC control. Although the seed-priming treatments modified these alterations in the SPM content in both genotypes, the responses depended on the Fusarium species. Under FV infection SAFV or PUFV treatments resulted in the lowest SPM contents among the treatments in both genotypes. At the same time, in the case of FG, DW and SA did not have influence but PUT pre-treatment increased the SPM levels compared to the only ABFG-treated samples in both genotypes and resulted in similar values as found in the ABC samples (Fig. 2C).
Comparing these changes with that of DAP (Fig. 2D), it can be concluded that in the susceptible line (EXPMV01), no remarkable changes were observed in the DAP amount after seed-priming treatment under control conditions (ABC, DWC, PUC, SAC) compared to the control (ABC). Althouhg after FV treatment (ABFV, DWFV, PUFV, SAFV), the seed soaking treatments decreased the levels of SPD and SPM, the DAP concentration did not increase. While after FG infection, in the susceptible genotype, the DAP level was almost doubled (ABFG) compared to the ABC, the DWFG, SAFG or PUFG pre-treatments alleviated this accumulation up to the lowest extent in the case of PUT priming (where the highest SPM accumulation was found). In contrast, in the tolerant genotype (EXPMV02), the seed soaking treatments significantly decreased the DAP accumulation after both Fusarium treatments. In addition, after FG infection, despite the observed SPD and SPM accumulation in PUT pre-treated samples, no increase was found in the DAP content compared to adequate control (ABFG) (Fig. 2D).
All three factors significantly influenced (p = 0.1%) the amounts of DAP, PUT, SPD and SPM measured in the coleoptile, although the results for DAP were affected by the genotypes × treatments only at p = 5%. Except for SPM, the results were mostly impacted by the genotypes. The amount of SPM was mostly influenced by the treatments, while the effects of the treatments on the other parameters had only secondary relevance. The combined effect of the two factors (genotypes × treatments) had the lowest influence on the measured values (Table 3).
Pathogen lifestyle influenced the polyamine content of the radicle more significantly than the genotype or seed-priming
As it was found in the coleoptile, similar general tendency in the changes of PA levels was observed in the radicle of the two genotypes (Fig. 3). Pronounced differences were detected rather between the FV- and FG-induced stress responses in both lines.
The FV infection (ABFV) alone increased the level of PUT (Fig. 3A) in both genotypes and the level of SPD (Fig. 3B) in the tolerant one (EXPMV02), while the concentration of SPM (Fig. 3C) decreased especially in the tolerant line compared to the absolute control (ABC). The DAP content did not change significantly in either genotype (Fig. 3D). Similarly, as it was described for the coleoptile, the FG infection induced more pronounced changes regardless of the degree of tolerance in both genotypes, namely high accumulation of PUT, but a decrease in SPD and SPM contents. The level of DAP increased only in the tolerant genotypes (EXPMV02) after FG infection compared to ABC.
Among the seed soaking treatments, under control conditions (DWC, SAC and PUC), the DW and PUT pre-treatments had similar effects, while the SA induced partly different changes in both lines. DW and PUT priming increased the level of PUT (Fig. 3A) in both genotypes but could increase the SPD level (Fig. 3B) only in the tolerant one (EXPMV02) compared to the control (ABC). The effect of the SA seed soaking was partly opposite: it only slightly raised the PUT content in the susceptible genotype (EXPMV01) but did not cause any changes in the tolerant one, whereas the SPD level lowered in the susceptible genotype and the tolerant one (EXPMV02) was not affected by the treatment compared to the ABC treatment. The SAC and PUC pre-treatments decreased the SPM content in the susceptible line (EXPMV01), while all the primings resulted in a decline for the tolerant line (Fig. 3C). The DAP amount did not change after the seed soaking treatments under control condition (Fig. 3D).
When seed soaking treatments were followed by FV infection, similar levels of PUT were detected to the corresponding control samples under control conditions (DWC, SAC and PUC), in both genotypes (Fig. 3A). Despite some statistically significant changes in the SPD content, the combined seed soaking and FV treatment did not induce remarkable changes either compared to adequate control (ABFV) (Fig. 3B). While the level of SPM was decreased by all the pre-treatments in both genotypes, and as the ABFV itself lowered the SPM concentration, the seed soaking treatments induced further drop in it (Fig. 3C). Among the pre-treatments only the DWFV seed soaking could induce a decrease in the DAP level during the FV infection for the tolerant genotype (EXPMV02) compared to the FV treatment alone (ABFV) (Fig. 3D).
When the FG treatment was followed the seed pre-treatments in the susceptible genotype (EXPMV01), the PUT and SPD content did not change, and the SPM content increased only in the case of the PUFG priming compared to adequte control (ABFG). The DAP content was decreased by DWFG and SAFG pre-treatments compared to that of the FG treatment alone (ABFG) in the susceptible genotype. In the tolerant line, (EXPMV02) SAFG and PUFG decreased the PUT level, DWFG increased the SPD level, SAFG and PUFG increased the SPM level, and all the pre-treatments decreased the DAP content compared to that of the ABFG-treated one (Fig. 3A–D).
The amounts of DAP, PUT, SPD and SPM measured in the radicle were significantly affected by all three factors (p = 0.1%). The genotypes had the greatest influence on the results followed by the treatments and then the combined effect of the genotypes × treatments (Table 4).
Discussion
PAs can have adverse roles: on the one hand, they serve as nutrients from the host cell to support fungal growth; on the other hand, however, they can limit the fungal spread. It can also be concluded that their accumulation and oxidation or even the back-conversion leads to H2O2 production, which steps are, not equally intensive during plant responses induced by biotrophic or necrotrophic pathogens. These responses play a beneficial role in plant defence mechanisms against biotrophic pathogens, while help the spread of necrotrophic pathogens (Pál and Janda 2017). Therefore, the fungi-induced modulation of host cell PA metabolism may lead to significant changes in the host’s susceptibility to different pathogens. However, the role of PA metabolism in the tolerance of plants to pathogens with different lifestyles is still not clear. Based on this, in the present study, our aim was to reveal the influence of the pathogenic lifestyle of Fusarium infection on the PA contents in two maize genotypes with different degree of tolerance. Particular attention was paid to investigate the effects of SA and PUT seed-priming on the tolerance manifested in the biomass parameters of the seedlings. In addition, it was also examined how these seed soaking pre-treatments influenced the PA metabolism related to the putative protective effect.
The infection with the hemibiotrophic FV in itself did not induce pronounced changes in the PA pool, except for a decrease in the SPM content in both the coleoptile and radicle. As the changes in PUT content were smaller than that of the SPD and SPM, the dominant role of the back-conversion of SPD/SPM to PUT could be excluded. Thus, the lowered level of higher PAs may be explained by their decreased synthesis, indicating that the inhibition of PA synthesis can be an important element in the defence mechanism against the hemibiotrophic fungi in both genotypes. Rodríguez-Kessler et al. (2008) analysed changes in the polyamine metabolism of maize in tumours formed during interaction with the biotrophic pathogenic fungus Ustilago maydis. In the tumour zone, 2:1 ratio of conjugated to free PUT was observed. These data suggest that the accumulation of conjugated PUT may play a dual role in the pathosystem of U. maydis. On the one hand, it promotes the accumulation of an antifungal agent involved in defence responses. On the other hand, it serves as a mechanism to reduce the level of free PAs that are prevalent in tumour growth. Investigation on the infection of tobacco with two biotrophic (Peronospora tabacina, Erysiphe cichoracearum) and a saprotrophic fungus (Alternaria tenuis), a bacterium (Pseudomonas tabaci) and a virus (TMV) Edreva (1997), revealed a reduction in the PA content of the infected tobacco leaves was observed compared to the control treatment. It was concluded that the extent of the reduction did not depend on the type of pathogens, but on the severity of the damage. It was also found that the decrease in PA content could be considered as a general response of tobacco to pathogen damage. In a recent review, it as summarised the role of PAs in plant disease resistance, with highlighting, among others, the role of H2O2 as a signalling molecule produced during the oxidation of PAs in addition to DAP accumulation (Takahashi 2016). In the present study the FG treatment itself, parallel with the decrease in SPD and SPM, induced DAP accumulation and a slight increase in PUT contents in both genotypes. These results confirmed that FG induced a more intensive terminal catabolism during the necrotrophic infection, which in turn increased the level of H2O2. Nevertheless, the increment in PUT level can be explained by the back-conversion or the decreased synthesis of higher polyamines or the induced PUT synthesis. Gardiner et al. (2010) also investigated the relationship between Fusarium graminearum infection and PAs. In their studies, PA content increased in infected plants and it was found that the increased PUT content may contribute to DON toxin production. In an investigation of the relationship between the also necrotrophic Aspergillus flavus infection and PA content, the high SPD/PUT ratio of the resistant lines compared to the susceptible ones suggested a possible role for host SPD in resistance to A. flavus aflatoxin production (Majumdar et al. 2019). Namely, a higher SPD content contributes to its greater availability for catabolism, conjugation and other regulatory roles. Overall, based on our results, it can be concluded that although there were some initial differences in the PA contents between the two genotypes, the degree of tolerance to Fusarium infection is not related to the changes in the PA contents, as FV or FG induced similar changes both in the tolerant (EXPMV02) and susceptible genotypes (EXPMV01), respectively. Montilla-Bascón et al. (2014) tested susceptible and resistant oat cultivars to biotrophic powdery mildew (Blumeria graminis f. sp. avenae) and found higher levels of PUT in susceptible cultivars than in resistant cultivars, while SPD content was higher in resistant cultivars than in susceptible ones. Among susceptible and resistant maize lines infected with necrotrophic Aspergillus flavus, higher levels of SPD and SPM were measured in resistant lines (Majumdar et al. 2019). In the present study, however, remarkable difference could be detected between the PA metabolism responses induced by the two Fusarium pathogens. DAP accumulation was induced only under FG infection, which was accompanied by a decrease in higher PA contents. This means that during the breakdown of SPD and SPM, a large amount of H2O2 can also accumulate in addition to DAP, which in turn can promote the spread of a necrotrophic fungus.
Based on the FW values, seed soaking treatments were beneficial against FV infection in both lines, while it had positive effects against FG infection only in the case of the tolerant genotype (EXPMV02). The biostimulating effect of SA and PUT seed soaking was also revealed in maize and wheat under normal growth conditions (Szalai et al. 2016; Tajti et al. 2018). In both cases, the positive effect was accompanied by the beneficial increment of PA contents. The two genotypes mostly responded similarly to the pre-treatments, but seed-primings with DW, SA or PUT had different effects on the PA levels under control condition and when it was followed by Fusarium infection. Remarkable differences were observed after SA seed soaking during the FG infection, as the PUT content increased dramatically in the susceptible genotype (EXPMV01) but it did not change in the tolerant one (EXPMV02) compared to the adequate SAC samples. Interestingly, an opposite tendency was observed for the SPD content also after SA pre-treatment under FG infection, as in the susceptible line (EXPMV01) it was decreased, but in the tolerant one (EXPMV02), it was increased. In the tolerant genotype (EXPMV02), under control condition, the seed soaking treatments already decreased the SPD and SPM content, which was not followed by the increase in PUT or DAP content. This means that neither the catabolism nor the back-conversion were activated, but the synthesis of the higher PAs was inhibited. The FV itself decreased the SPM level, and when it was preceded by seed soaking, the SPD and SPM contents decreased, but the DAP level, too. During the FG infection, the accumulation of DAP was reduced by the seed soaking treatments, especially SA and PUT pre-treatments most pronouncedly in the coleoptile in both genotypes. Moreover, PUT seed soaking increased the amount of SPM in the susceptible (EXPMV01) and the tolerant lines (EXPMV02). Therefore, the increased total PA amount with the decreased catabolism can be responsible for the beneficial effect of the seed soaking against the necrotrophic pathogen. Nevertheless, in the coleoptile of the tolerant genotype (EXPMV02), seed soaking treatments were also able to reduce DAP accumulation during FV infection, even though FV alone did not increase its amount.
Conclusions for future biology
PAs have important role in the biotic stress responses of plants. Plant tissues respond rapidly to the infection with pathogens, and the responses can depend on the lifestyle of the pathogen. Accumulation of PUT with the decrease in the SPD/SPM content, and in turn, their decreased catabolism may inhibit the necrotrophic pathogen. At the same time, the accumulation of PA and their metabolism lead to increased H2O2 content, which prevents the spread of biotrophic pathogens. Fine-tuning of PA metabolism is important under either condition. Therefore, more precise information on the timing and location of changes in PA metabolism is needed for the understanding of PA-related resistance.
Application of plant hormones, like SA or biostimulants, such as PAs to seeds is an environmental-friendly technique in crop protection, and may protect plants from an early age. Seed-priming can induce physiological, transcriptional, metabolic, and epigenetic changes and effectively enhance disease resistance. However, before it can be commercially used, several aspects should be elucidated. It is justified, among others, to investigate the duration of the priming effect and the range of biotic stresses.
References
Alzahrani Y, Rad MM (2019) Compared to antioxidants and polyamines, tre role of maize grain-derived organic biostimulants in improving cadmium tolerance in wheat plants. Ecotoxicol Environ Saf 182:109378. https://doi.org/10.1016/j.ecoenv.2019.109378
Amborabé BE, Fleurat-Lessard P, Chollet JF, Roblin G (2002) Antifungal effects of salicylic acid and other benzoic acid derivatives towards Eutypa lata: structure-activity relationship. Plant Physiol Biochem 40(12):1051–1060
Beccaccioli M, Salustri M, Scala V, Ludovici M, Cacciotti A, D’Angeli S, Brown DW, Reverberi M (2021) The effect of Fusarium verticillioides fumonisins on fatty acids, sphingolipids, and oxylipins in maize germlings. Int J Mol Sci 22(5):2435. https://doi.org/10.3390/ijms22052435
Canales FJ, Montilla-Bascón G, Rispail N, Prats E (2019) Salicylic acid regulates polyamine biosynthesis during drought responses in oat. Plant Signal Behav 14(10):e1651183. https://doi.org/10.1080/15592324.2019.1651183
Cvikrová M, Gemperlová L, Martincová O, Radomira V (2013) Effect of drough and combined drough and heat stress on polyamine metabolism in proline- over- producing tobacco plants. Plant Physiol Biochem 73:7–15. https://doi.org/10.1016/j.plaphy.2013.08.005
Daniels BA (1983) Elimination of Fusarium moniliforme from corn seed. Plant Dis 67(6):609–611
Ding Y, Sun T, Ao K, Peng Y, Zhang Y, Li X, Zhang Y (2018) Opposite roles of salicylic acid receptors NPR1 and NPR3/NPR4 in transcriptional regulation of plant immunity. Cell 173(6):1454–1467. https://doi.org/10.1016/j.cell.2018.03.044
Doohan FM, Brennan J, Cooke BM (2003) Influence of climatic factors on Fusarium species pathogenic to cereals. Eur J Plant Pathol 109:755–768. https://doi.org/10.1007/978-94-017-1452-5_10
Edreva A (1997) Tobacco polyamines as affected by stresses induced by different pathogens. Biol Plant 40:317–320
Farooq M, Basra SMA, Rehman H, Hussain M (2008) Seed priming with polyamines improves the germination and early seedling growth in fine rice. J New Seeds 9(2):145–155. https://doi.org/10.1080/15228860802087297
Folcher L, Jarry M, Weissenberger A, Geraul F, Eychenne N, Delos M, Regnault-Roger C (2009) Comparative activity of agrochemical treatments on mycotoxin levels with regard to corn borers and Fusarium mycoflora in maize (Zea mays L.) fields. Crop Prot 28(4):302−308 doi.org: https://doi.org/10.1016/j.cropro.2008.11.007
Gao Q-M, Zhu S, Kachroo P, Kachroo A (2015) Signal regulators of systemic acquired resistance. Front Plant Sci 6:228. https://doi.org/10.3389/fpls.2015.00228
Gardiner MD, Kazan K, Praud S, Torney FJ, Rusu A, Manners MJ (2010) Early activation of wheat polyamine biosynthesis during Fusarium head blight implicates putrescine as an inducer of trichothecene mycotoxin production. Plant Biol 10:289. https://doi.org/10.1186/1471-2229-10-289
Gill SS, Tuteja N (2010) Polyamines and abiotic stress tolerance in plants. Plant Signal Behav 5(1):26–33. https://doi.org/10.4161/psb.5.1.10291
Guenther JC, Trail F (2005) The development and differentiation of Gibberella zeae (anamorph: Fusarium graminearum) during colonization of wheat. Mycologia 97(1):229–237. https://doi.org/10.1080/15572536.2006.11832856
Handa AK, Nambeesan S, Mengiste T, Laluk K, Abu Qamar S, Mattoo AK (2011) Polyamine spermidine is an upstream negator of ethylene-regulated pathogenesis of Botrytis Cinerea in tomato leaf. Acta Hortic 914:109–112. https://doi.org/10.17660/ActaHortic.2011.914.18
Harris LJ, Balcerzak M, Johston A, Schneiderman D, Ouellet T (2016) Host-preferential Fusarium graminearum gene expression during infection of wheat, barley and maize. Fungal Biol 120(1):11–123. https://doi.org/10.1016/j.funbio.2015.10.010
Hasanuzzaman M, Alhaithloul HAS, Parvin K, Borhannuddin Bhuyan MHM, Tanveer M, Mohsin SM, Nahar K, Soliman MH, Mahmud JA, Fujita M (2019) Polyamine action under metal/metalloid stress: regulation of biosynthesis, metabolism, and molecular interactions. Int J Mol Sci 20(13):3215. https://doi.org/10.3390/ijms20133215
Hussain SS, Ali M, Ahmad M, Siddique MHK (2011) Polyamines: natural and engineered abiotic and biotic stress tolerance in plants. Biotechnol Adv 29(3):300–311. https://doi.org/10.1016/j.biotechadv.2011.01.003
Jacob D, Petersen J, Eggert B, Alias A, Christensen OB, Bouwer LM et al (2014) Euro-cordex: new high-resolution climate change projections for European impact research. Reg Environ Change 14(2):562–578. https://doi.org/10.1007/s10113-013-0499-2
Janse van Rensburg HC, Limami AM, Van den Ende W (2021) Spermine and spermidine priming against Botrytis cinerea modulates ROS dynamics and metabolism in Arabidopsis. Biomolecules 11(2):223. https://doi.org/10.3390/biom11020223
Jiménez-Bremont JF, Ruiz OA, Rodríguez-Kessler M (2007) Modulation of spermidine and spermine levels in maize seedlings subjected to long-term salt stress. Plant Physiol Biochem 45(10–11):812–821. https://doi.org/10.1016/j.plaphy.2007.08.001
Leslie JF and Summerell BA (2006) The Fusarium Laboratory Manual. Blackwell Publishing (eds: Leslie JF.and Summerell BA) 388 p. DOI: https://doi.org/10.1002/9780470278376
Li L, Zhu T, Song Y, Feng L, Kear PJ, Riseh RS, Sitohy M, Datla R, Ren M (2022) Salicylic acid fights against Fusarium wilt by inhibiting target of rapamycin signaling pathway in Fusarium oxysporum. J Adv Res 39:1–13
Maiorani A, Reyneri A, Sacco D, Magni A, Ramponi C (2008) A dynamic risk assesment model (FUMAgrain) of fumonisin synthesis by Fusarium verticillioides in maize grain in Italy. Crop Prot 28(3):243–256. https://doi.org/10.1016/j.cropro.2008.10.012
Majumdar R, Minocha R, Lebar MD, Rajasekaran K, Long S, Carter-Wientjes C, Minocha S, Cary JW (2019) Contribution of maize polyamine and amino acid metabolism toward resistance against Aspergillus flavus infection and aflatoxin production. Front Plant Sci 10:692. https://doi.org/10.3389/fpls.2019.00692
Meissle M, Mouron P, Musa T, Bigler F, Pons X, Vasileiadis VP, Otto S, Antichi D, Kiss J, Palinkas Z, Dorner Z, Van Der Weide R, Groten J, Czembor E, Adamczyk J, Thibord JB, Melander B, Nielsen GC, Poulsen RT, Zimmermann O, Verschwele A, Oldenburg E (2010) Pests, pesticide use and alternative options in European maize production: current status and future prospects. J Appl Entom 134:357–375. https://doi.org/10.1111/j.1439-0418.2009.01491.x
Mesterházy Á, Lememns M, Reid LM (2011) Breeding for resistance to ear rots caused by Fusarium spp. in maize. Plant Breed 131:1–19. https://doi.org/10.1111/j.1439-0523.2011.01936.x
Montilla-Bascón G, Rubiales D, Prats E (2014) Changes in polyamine profile in host and non-host oat–powdery mildew interactions. Phytochem Lett 8:207–212. https://doi.org/10.1016/j.phytol.2014.01.002
Moschou PN, Sarris PF, Skandalis N, Andriopoulou AH, Paschalidis KA, Panopoulos NJ, Roubelakis-Angelakis KA (2009) Engineered polyamine catabolism preinduces tolerance of tobacco to bacteria and oomycetes. Plant Physiol 149(4):1970–1981. https://doi.org/10.1104/pp.108.134932
Munkvold GP (2003) Epidemiology of Fusarium diseases and their mycotoxins in maize ears. Eur J Plant Pathol 109:705–713. https://doi.org/10.1023/A:1026078324268
Németh M, Janda T, Horváth E, Páldi E, Szalai G (2002) Exogenous salicylic acid increases polyamine content but may decrease drought tolerance in maize. Plant Sci 162(4):569–574. https://doi.org/10.1016/S0168-9452(01)00593-3
Nyvall RF, Kommedahl T (1970) Saprophytism and survival of Fusarium moniliforme in corn stalks. Phytopathology 60:1233–1235
Pál M, Janda T (2017) Role of polyamine metabolism in plant pathogen interactions. J Plant Sci Phytopathol 1:95–100. https://doi.org/10.29328/journal.jpsp.1001012
Pál M, Szalai G, Janda T (2015) Speculation: polyamines are important in abiotic stress signaling. Plant Sci 237:16–23. https://doi.org/10.1016/j.plantsci.2015.05.003
Pál M, Szalai G, Gondor OK, Janda T (2021) Unfinished story of polyamines: role of conjugation, transport and light-related regulation in the polyamine metabolism in plants. Plant Sci 308:110923. https://doi.org/10.1016/j.plantsci.2021.110923
Qi PF, Johnston A, Balcerzak M, Rocheleau H, Harris LJ, Long XY, Wei YM, Zheng YL, Ouellet T (2012) Effect of salicylic acid on Fusarium graminearum, the major causal agent of fusarium head blight in wheat. Fungal Biol 116(3):413–426. https://doi.org/10.1016/j.funbio.2012.01.001
Quarantin A, Hadeler B, Kröger C, Schäfer W, Favaron F, Sella L, Martínez-Rocha AL (2019) Different hydrophobins of Fusarium graminearum are involved in hyphal growth, attachment, water-air interface penetration and plant infection. Front Microbiol 10:751. https://doi.org/10.3389/fmicb.2019.00751
Radojicic A, Li X, Zhang Y (2018) Salicylic acid: a double-edged sword for programmed cell death in plants. Front Plant Sci 9:1133. https://doi.org/10.3389/fpls.2018.01133
Ramazan S, Nazir I, Yousuf W, John R (2022) Environmental stress tolerance in maize (Zea mays): role of polyamine metabolism. Funct Plant Biol 50(2):85–96. https://doi.org/10.1071/FP21324
Rodríguez-Kessler M, Ruiz OA, Maiale S, Ruiz-Herrera J, Jiménez-Bremont JF (2008) Polyamine metabolism in maize tumors induced ba Ustilago maydis. Plant Physiol Biochem 46(8–9):805–814. https://doi.org/10.1016/j.plaphy.2008.05.012
Sadeghipour O (2019) Polyamines protect mung bean Vigna radiata (L.) Wilczek plants against drought stress. Biol Fut 70(1):71–78
Sutton JC (1982) Epidemiology of wheat head blight and maize ear rot caused by Fusarium graminearum. Can J Plant Pathol 4:195–209. https://doi.org/10.1080/07060668209501326
Sváb J (1981) Biometriai módszerek a mezőgazdasági kutatásban. Mezőgazdasági Kiadó, Budapest, 490 p.
Szalai G, Pál M, Árendás T, Janda T (2016) Priming seed with salicylic acid increases grain yield and modifies polyamine levels in maize. Cereal Res Commun 44:537–548. https://doi.org/10.1556/0806.44.2016.038
Szepesi Á, Poór P, Bakacsy L (2022) Short-term salicylic acid treatment affects polyamine metabolism causing ROS–NO imbalance in tomato roots. Plants 11(13):1670. https://doi.org/10.3390/plants11131670
Szőke C, Nagy Z, Gierczik K, Székely A, Spitkó T, Tóth Zsubori Z, Galiba G, Marton CL, Kutasi K (2018) Effect of the afterglows of low pressure Ar/N2 −O2 surface-wave microwave discharges on barley and maize seeds. Plasma Process Polym 15:e1700138. https://doi.org/10.1002/ppap.201700138
Szőke C, Pál M, Szalai G, Janda T, Rácz F, Marton LC (2011) Potencial role of salicylic acid in tolerance of maize to Fusarium graminearum. Acta Biol Szeged 55(1):167−168. https://abs.bibl.u-szeged.hu/index.php/abs/article/view/2741
Tajti J, Janda T, Majláth I, Szalai G, Pál M (2018) Comparative study on the effects of putrescine and spermidine pre-treatment on cadmium stress in wheat. Ecotoxicol Environ Saf 148:546–554. https://doi.org/10.1016/j.ecoenv.2017.10.068
Takahashi Y (2016) The role of polyamines in plant diseaes resistance. Environ Control Biol 54(1):17–21. https://doi.org/10.2525/ecb.54.17
Vigier B, Reid LM, Seifert KA, Stewart DW, Hamilton RI (1997) Distribution and prediction of Fusarium species associated with maize ear rot in Ontario. Can J Plant Pathol 19(1):60–65. https://doi.org/10.1080/07060669709500574
Wang W, Paschalidis K, Feng JC, Song J, Liu JH (2019) Polyamine catabolism in plants: a universal process with diverse functions. Front Plant Sci 10:561
Yu Z, Jia D, Liu T (2019) Polyamine oxidases play various roles in plant development and abiotic stress tolerance. Plants 8(6):184. https://doi.org/10.3390/plants8060184
Funding
Open access funding provided by ELKH Centre for Agricultural Research. This work was financed by grants from the National Research Development and Innovation Office, Ministry of Innovation and Technology of Hungary from the National Research, Development and Innovation Fund, Hungary (NKFIH K134395 and TKP2021-NKTA).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kovács, B., Kovács, A., Pál, M. et al. Changes in polyamine contents during Fusarium graminearum and Fusarium verticillioides inoculation in maize seedlings with or without seed-priming. BIOLOGIA FUTURA 74, 145–157 (2023). https://doi.org/10.1007/s42977-023-00162-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42977-023-00162-7