
Abstract
The Aysén-Río Mayo Basin was a back-arc/marginal basin developed in southwestern South America (43°–47°S) between the Tithonian–Aptian. Its sedimentary fill corresponds to the Coyhaique Group, which represents a transgressive–regressive succession. Six lithofacies and five microfacies were defined for three outcrops exposed south of Coyhaique (45°40’S). The outcrops have a mixed calcareous–volcaniclastic composition and were assigned to the early transgressive Toqui Formation, i.e., lowermost part of the Coyhaique Group. These mixed rocks comprise bioclastic–volcaniclastic conglomerate, gravelly allochemic sandstone, and gravelly–sandy allochem limestone. Bedding is sharp to amalgamated, sometimes rippled, depicting a wave- and storm-influenced, mixed inner- to mid-ramp. The ramp developed over a Valanginian, active volcanic terrain (Foitzick Volcanic Complex), source of the volcaniclastic sediments. Limestones are rich in reworked bioclasts, and controlled by calcitic organisms including gryphaeid oysters, non-geniculate red algae, and echinoid fragments, defining a heterozoan association (“maerl”-like sediments); less frequent are ahermatypic corals, serpulids, and carbonized wood. Based on their inferred paleolatitude (south of 45°–50°S), fossil assemblage (heterozoan), and kind of carbonate platform (ramp-type), these calcareous rocks of the Toqui Formation depict a “cool-water” (sensu lato), non-tropical setting. The fossil assemblage includes oysters (Aetostreon spp.), and abundant calcareous red algae attributed to Archamphiroa jurassica Steinmann (1930), a taxon previously known from the upper Tithonian Cotidiano Formation of Argentina. A. jurassica is here reported for the first time for the Lower Cretaceous of Chile, suggesting a broader upper Tithonian—Valanginian-Hauterivian? range for the species. The facies model presented here contrasts with the depositional environments depicted for correlative reefal rocks in Argentina (Tres Lagunas Formation), which reflect a “warm-water” setting. In the Aysén-Río Mayo Basin, the influence of sea-water key physical variables in the carbonate sedimentation, as well as the position and hydraulic regime of the carbonate platforms within the basin, and their interaction with the volcanism are still unclear.
Similar content being viewed by others


Introduction
Mixed calcareous–volcaniclastic rocks are common constituents in the Late Jurassic–Early Cretaceous of southwestern South America (Central Patagonia, 43°–47°S); these deposits represent the early transgressive phase in the Aysén—Río Mayo Basin, whose marine stage developed during the Tithonian–Aptian (e.g., Aguirre-Urreta and Ramos 1981; Ramos 1981; Scasso 1987; 1989; Bell et al. 1996; Folguera and Iannizzotto 2004; Suárez et al. 2010a, b; Fig. 1A, B). In Chile, these mixed rocks are exposed as scattered outcrops in the Los Lagos and Aysén regions (De la Cruz et al. 1996; Welkner et al., 2004) and have been referred to as the Toqui Formation (Bell et al. 1994; Suárez and De la Cruz 1994a,b; Fig. 1A, B). This unit comprises two members (sensu Rivas et al. 2021): the mixed, calcareous–volcaniclastic Manto Member conforming the basal portion of the Toqui Fm., and the volcaniclastic San Antonio Member which overlies and interfingers the Manto Member. Both members display marked lateral- and latitudinal facial changes, reflecting strong diachronism during the Tithonian–Valanginian interval (Suárez et al. 2005; 2010a, b), and the development of mixed, shallow-marine platforms over an irregular extensional topography (Scasso 1989; Hechem et al. 1993; Folguera and Iannizzotto 2004; Rivas et al. 2021).

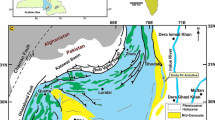
Stratigraphy and location. A Regional chronostratigraphic chart (modified from Rivas et al. 2021). Some formational names have been abbreviated: Fm Formation, APB Arroyo Pedregoso Beds, ABB Arroyo Blanco Beds, BNVC Baño Nuevo Volcanic Complex, FVC Foitzick Volcanic Complex, TLF Tres Lagunas Formation (see “Stratigraphy and correlations”). B Regional map displaying the location of the Aysén-Río Mayo Basin, the Austral Basin, and localities mentioned in the text. Aysén-Río Mayo basin: dotted outline sensu Aguirre-Urreta and Ramos (1981); dashed outline sensu Suárez et al. (2010a); Austral Basin outline sensu Cuitiño et al. (2019). C Local geological map of the La Plata Lake and Coyhaique areas (simplified from Lizuaín et al. 1995; De la Cruz et al. 2003; Suárez et al. 2007). Given the small areal exposures of the early transgressive, mixed- and calcareous rocks, all the formations are represented as part of the Coyhaique Group (undifferentiated). D Study area: Close-up of the area south of Coyhaique, displaying the outcrops studied here (modified from De la Cruz et al. 2003). ab Arroyo Blanco, ac Arroyo Cotidiano (type locality of the Cotidiano Fm.), ap Arroyo Pedregoso, eh Estero La Horqueta, gl General Carrera/Buenos Aires Lake, pa Palena (town), R7 Route 7 (Carretera Austral), tl Tres Lagunas (Laguna Salada, type locality of the Tres Lagunas Fm.), tm El Toqui Mine (type locality of the Toqui Fm.). Outcrops here addressed: MCH Muralla China, LRO La Rosita, SRP2 Salto Río Pollux
In Argentina, the Toqui Formation has been correlated with coeval (Tithonian–Valanginian), calcareous and mixed units exposed in the area adjacent to the La Plata—Fontana lakes (e.g., Cotidiano and Tres Lagunas Formations; Sect. “Stratigraphy and correlations”; Fig. 1A). Overall, these mixed rocks represent transgressive deposits, depicted as settled in euhaline, shallow marine environments, and locally influenced by active volcanism (e.g., Ramos 1981; Ploszkiewicz and Ramos 1977; Olivero 1982; Scasso 1987; 1989; Bell et al. 1996; Suárez et al. 1996; De la Cruz et al. 2003; Scasso and Kiessling 2002; Iannizzotto et al. 2004).
Given the discrete, discontinuous, and diachronic exposure of these mixed calcareous–volcaniclastic units, they have been studied and interpreted separately (see review in Rivas et al. 2021; Fig. 1C), and several regional models for its sedimentation have been proposed (e.g., Skarmeta 1976; Aguirre-Urreta and Ramos 1981; Scasso 1989; Hechem et al. 1993; Bell et al. 1994; Townsend 1998; Folguera and Iannizzotto 2004; Suárez and De la Cruz 1994a, b). Here, a sedimentological and microfacial analysis is presented of three mixed volcaniclastic-calcareous sections, assigned to the Toqui Formation, and exposed south of Coyhaique in southern Chile (45°40°S; Fig. 1D). This study improves the understanding of the depositional environments of mixed rocks included in the Toqui Formation, and their relationship with the Argentinian exposures is also discussed. The outcrops presented here provide new insight into non-tropical or “cool-water” carbonate sedimentation as well as the development of carbonate ramps in volcanic arc settings.
Geological setting
The Aysén—Río Mayo Basin was a back-arc/marginal basin (see models without seafloor-spreading in Tarney et al. 1981; Artemieva 2023) formed at the southwestern margin of West Gondwana (proto-South America, ca. 43°–49°S) during the Late Jurassic and Early Cretaceous (Tithonian—Aptian, e.g., Aguirre-Urreta and Ramos 1981; Suárez et al. 2010a, b; Fig. 1B). Its onset is regarded as triggered by back-arc extension to the east of the coeval volcanic arc (Suárez et al. 2010a, b; Echaurren et al. 2016), which is represented by the Patagonian Batholith (Pankhurst et al. 1999; Suárez and De la Cruz 2001; Suárez et al. 2023; Fig. 1C). The basin developed unconformably over the basement, which consists of upper Paleozoic metamorphic and metasedimentary rocks as well as Lower Jurassic sedimentary units (e.g., Olivero 1982; Ploszkiewicz 1987; De la Cruz et al. 2003; Hervé et al. 2008; Fig. 1A).
The oldest units conforming the Aysén—Río Mayo basin fill, correspond to the Ibáñez Formation in Chile and the correlative Lago La Plata Formation in Argentina (e.g., Ramos 1976; Haller et al. 1981; Haller and Lapido 1982; De la Cruz et al. 1994; 2003; Fig. 1A). The Ibáñez Formation consists of calc-alkaline, subaerial- and minor submarine-settled, felsic volcanic- and volcaniclastic rocks, depicting the Late Jurassic–Early Cretaceous (Oxfordian?—early Hauterivian) extrusive counterpart of the Patagonian Batholith (Pankhurst et al. 2003; Suárez et al. 2005). These volcanic rocks syn-tectonically filled the extensional topography (Echaurren et al. 2016; Folguera and Iannizzotto 2004), whereas its uppermost strata underlie and interfinger with the shallow marine deposits of the Toqui Formation and correlative units (see Sect. “Stratigraphy and correlations” below), i.e., the lowermost part of the Coyhaique Group (e.g., Skarmeta 1976; Olivero 1982; Scasso 1989; Covacevich et al. 1994; Suárez et al. 2009; 2010a; Fig. 1A).
The Coyhaique Group is a transgressive–regressive succession of Tithonian–Aptian age (Ramos 1981; Scasso 1987; Covacevich et al. 1994; Bell et al. 1994; Suárez et al. 2010a; Suárez and De la Cruz 1994a, b), and linked to a back-arc “Pacific” marine incursion caused by tectonic-thermal subsidence (Skarmeta 1976; Haller and Lapido 1980; Aguirre-Urreta and Ramos 1981). In Chile, the Coyhaique Group includes three formations (Fig. 1A): (1) the Toqui Formation (Tithonian—Valanginian, Hauterivian?), corresponding to shallow-marine, mixed (calcareous–volcaniclastic) and tuffaceous rocks, deposited during the early transgression (De la Cruz et al. 1994; Suárez et al. 2009; Rivas et al. 2021); (2) the Katterfeld Formation (Valanginian-Barremian; Masiuk and Nakayama 1978; Aguirre-Urreta et al. 2000; Olivero and Aguirre-Urreta 2002; Kesjár et al. 2017) overlies the Toqui Fm. with a transitional or conformable contact and consists of tuffaceous and carbonaceous black mudstone, settled in offshore environments during the post-rift stage (Scasso 1989; Suárez et al. 2007; 2010a); and 3) the Apeleg Formation (Hauterivian-Aptian) overlies the Katterfeld Formation with a gradational contact; it includes heterolithic, sandy, and gravelly facies, depicting tidal, and deltaic-paralic, regressive successions (e.g., Ramos 1981; Scasso 1989; Gonzalez-Bonorino and Suárez 1995; Bell and Suárez 1997; Fig. 1A).
The Aysén-Río Mayo Basin was inverted during the Aptian, linked to compression after a fast, westwards relative movement of the South American Plate (Suárez et al. 2010a, b; Gianni et al. 2019; Ramos et al. 2019). The latter reactivated the volcanic arc, resulting in local Surtseyan eruptions (Baño Nuevo Volcanic Complex; Demant et al. 2010; Suárez et al. 2010b; Fig. 1A), and the effusion of subaerial, intermediate, volcanic rocks of the Divisadero Group (Aptian—early Cenomanian). The Divisadero Group covers the underlying Coyhaique Group with a unconformable, but locally (para-)conformable contact (e.g., Aguirre-Urreta and Ramos 1981; Ramos 1981; De la Cruz et al. 1994; Suárez et al. 2010a; Fig. 1A).
Stratigraphy and correlations
In central Patagonia (43°–47°S), the early transgressive deposits of the Aysén-Río Mayo Basin (ARB) comprise a Tithonian – Hauterivian? range (e.g., Haller and Lapido 1980; Olivero 1983; 1987; Scasso 1989; Covacevich et al. 1994; Suárez et al. 2009), reflecting strong diachronism during the transgression (Suárez et al. 2005; Fig. 1A). The early transgressive rocks of the ARB have a mixed- and calcareous composition, and they depict shallow marine settings near volcanic centers (e.g., Ramos 1978; Suárez et al. 1996, 2009; Scasso and Kiessling 2002; Ramos and Lazo 2006; Rivas et al. 2021, and references therein).
In Argentina, these rocks have been treated as singular units (Fig. 1A), including: (i) the Cotidiano Formation (Oxfordian-Kimmeridgian sensu Ramos 1976; Ramos 1981; upper Tithonian sensu Olivero 1987; Bucur et al. 2009); (ii) the “Arroyo Pedregoso Beds” (upper Tithonian sensu Ramos 1981; Olivero 1982; 1987); (iii) the “Arroyo Blanco Beds” (Berriasian-Hauterivian sensu Olivero 1982; 1987; upper Valanginian sensu Olivero and Aguirre-Urreta 2002; Fig. 1A); and (iv) the Tres Lagunas Formation (at its type locality “Laguna Salada”, Valanginian-lower Hauterivian sensu Ploszkiewicz and Ramos 1977; Berriasian sensu Olivero 1983; Tithonian?—upper Valanginian sensu Scasso and Kiessling 2002; Fig. 1A).
The former three were regarded by Olivero (1987) as sedimentary alternations within the Lago la Plata Formation; this approach was followed by Covacevich et al. (1994) for some calcareous outcrops exposed north of the town of Ñireguao, including them within the Ibáñez Formation. In the Chilean side, these mixed- and calcareous outcrops were later grouped together in the Toqui Formation (e.g., Bell et al. 1994; De la Cruz et al. 1996; 2003; Suárez and De la Cruz 1994a, b). The Toqui Formation was redefined by Rivas et al. (2021; after Bell et al. 1994; Suárez et al. 1996; Suárez and De la Cruz 1994a, b), as conformed by two members: the mixed calcareous–volcaniclastic Manto Member, interbedded and overlied by the volcaniclastic San Antonio Member. In the area south of Coyhaique (45°35’S; Fig. 1D), these members appear discretely as mixed, either calcareous- or volcaniclastic-rich outcrops, depicting an upper Berriasian/lower Valanginian – Hauterivian? range (see “Age of the deposits”).
Age of the deposits
In the study area, the age of the Toqui Formation is interpreted to be late Berriasian/early Valanginian-Hauterivian? South of “Laguna Foitzick”, a late Berriasian age is based on a faunal association including Neocomitidae indet., and the similarity of this fauna with Berriasian outcrops exposed north of Ñireguao, at the “Estero La Horqueta” (Covacevich et al. 1994; Suárez et al. 1996; De la Cruz et al. 2003; Fig. 1C). In Laguna Foitzick, the Toqui Formation overlies the Foitzick Volcanic Complex, a local member of the Ibáñez Formation, with an erosion-angular unconformity, but locally displaying a peperitic contact (Bell et al. 1994; Suárez et al. 1996; Suárez and De la Cruz 1994a, b; Fig. 1A). There, rocks from the uppermost volcanic strata, below the unconformity, were dated to U–Pb 138.3 ± 1.3 Ma (Pankhurst et al. 2003), providing a “Valanginian or even younger” lower range limit for the sedimentary succession in the Coyhaique area (Suárez et al. 2005). A broader Berriasian-Hauterivian age (“Neocomian”) was inferred by Rubilar (2000) near the “Salto del Pollux”, based on a tentative new species of Aetostreon sp. In the same locality, Cecioni and Charrier (1974) reported findings of “Belemnopsis patagoniensis”, commonly found in the Lower Cretaceous of southernmost South America (Riccardi 1977; Aguirre-Urreta 2002; Ippolitov et al. 2015).
The top of the Toqui Formation is not exposed in the study area; however, in its type locality (El Toqui Mine), it pass transitionally to tuffaceous- and black mudstone of the Katterfeld Formation (Bussey et al., 2010; Rivas et al. 2021). At the Salto del Pollux locality (outcrop SRP2; Fig. 1D), the Toqui Formation is in tectonic contact with black mudstone of the Katterfeld Formation. There, the marine succession is confined by a post-depositional basaltic andesite sill dated to Ar–Ar 61 ± 0.4 Ma (“Muralla China Sill”; Petford and Turner 1996; De la Cruz et al. 2003), intruding both the Toqui- and Katterfeld Formations (Fig. 1D). Regionally, the Katterfeld Formation provides a Valanginian—Barremian upper range limit for the Toqui Formation (Masiuk and Nakayama 1978; Aguirre-Urreta et al. 2000; Olivero and Aguirre-Urreta 2002; Kesjár et al. 2017).
Materials and methods
This study comprises three sedimentary sections exposed between south of the Laguna Foitzick and the waterfall of the Pollux River (known as “El Salto”, Fig. 1D), between 8 and 11 km south of Coyhaique, capital of the Aysén Region, Chile (45°34’S; Fig. 1B–C). They are accessed via Route 7. The three outcrops define a ca. 67 m-thick composite log (outcrops LRO, MCH, SRP2; Fig. 1D), and they have been assigned to the Toqui Formation (Suárez and De la Cruz 1994a, b; De la Cruz et al. 2003). Based on their lithology, these stratified rocks are here arranged in six lithofacies (Table 2); five microfacies were additionally defined based on the petrographic analysis of fourteen thin sections (Table 1). The outcrop LRO has been previously addressed (Bell et al. 1994; Townsend 1998; Suárez and De la Cruz 1994a, b) as well as outcrop SRP2 (Katz 1961).
Grain-size categories used here follow Blair and McPherson 1999, and modal compositions of sandstone are based on Garzanti 2019. Lithological description of limestone is after Wright (1992), whereas non-genetic schemes were preferred for mixed- (Mount 1985), and volcaniclastic sediments (Fisher 1961; 1966; Fisher and Schmincke 1984; Cas and Wright 1987). A brief glossary of the volcanic terms applied here can be found as supplementary material; the term “volcaniclastic” was preferred (instead of “volcanic”), to highlight the epiclastic nature of the volcanic material (e.g., volcaniclastic sandstone). Given the mixed nature of most of rocks analyzed here, depositional models for the interpretation of siliciclastic (e.g., Clifton 2006; Plint 2010), and for carbonate environments were required (e.g., Burchette and Wright 1992; Flügel 2010). Raw lithological data taken in the field are presented as supplementary material (Tables SM-1 to SM-3).
Measurements of microfossils were registered with the software ImageJ, using micro-photographs taken with an Olympus camera, model PEN Lite E-PL7. SEM images of thin sections were taken with a microscope ZEISS WITec RISE EVO MA15 at the Universität Heidelberg, following the procedure of Munnecke et al. (2000). Calculations were made in Microsoft Excel. Chronostratigraphic categories used here correspond with the ICS International Stratigraphic Chart v2022/02 (Cohen et al. 2013; updated). Illustrations were created with Adobe Illustrator CS6. Rocks and fossils samples are kept in the Geology Department of the Universidad Mayor in Santiago, Chile, as a temporary repository.
Facies analysis
The litho- and microfacies of the Toqui Formation analysed here have a mixed volcaniclastic–bioclastic composition. Rocks were classified into six lithofacies (see “Lithofacies”; Table 2), supported by the characterization of five microfacies (Table 1), presented below.
Petrography
Microfacies comprise both ends of mixed rocks (calcareous- and terrigenous-rich), varying between gravelly–sandy allochem limestone and gravelly allochemic sandstone (sensu Mount 1985). Calcareous-rich microfacies include volcaniclastic–bioclastic, pack–rudstone, float–wackestone, and grainstone (microfacies Mf-1 to Mf-3; Table 1, Figs. 2; 3). On the other hand, terrigenous-rich microfacies consist of bioclastic–volcaniclastic, quartzo-litho-feldspathic- to quartzo-feldspathic sandstone (microfacies Mf-4, Mf-5).

Petrography and microfacies (Mf). A-C: LRO outcrop (Sect. “La Rosita Section (LRO) - gravelly, mixed inner ramp”). A Sandy–gravelly, volcaniclastic–bioclastic pack–rudstone with wackestone patches (Mf-1). B Another view of the previous sample (Fig. 2A) showing reworked fossils (some with an incipient micritic coating) and cementation (microfacies Mf-1). C Gravelly, bioclastic–volcaniclastic, quartzo-feldspatho-lithic sandstone with pseudo-grainstone patches (granular cements; sub-type Mf-4c). D–F MCH outcrop (Sect. “Muralla China Section (MCH) - wave- and storm-dominated inner-ramp”). D Sandy–gravelly, volcaniclastic–bioclastic float–wackestone with fitted packstone patches (Mf-2); E Sandy bioclastic grainstone; F red algae-rich grainstone (A. jurassica). G–K SRP2 outcrop (Sect. “Salto Río Pollux Section (SRP2)—mixed, distal mid-ramp”). G Gravelly, muddy, bioclastic–volcaniclastic (quartzo) feldspatho-lithic sandstone (rich in organic-matter; Mf-4om); note pressure solution and differential alteration of volcanic lithics (rim-center). H Similar to previous, but finer-grained, poorer in organic matter and richer in argillaceous matrix. I Subtype of Mf-4 with poikilotopic calcareous cement (Mf-4c). J Alternation of (mixed) bioclastic-volcaniclastic, medium-coarse-grained, quartzo-feldspatho-lithic sandstone (Mf-4) and fine-medium-grained, litho-quartzo-feldspathic sandstone (Mf-5); dotted line marks the contact; note parallel lamination (micas). K Close-up of Mf-5 displaying its volcaniclastic-rich composition, and its strong silicification; dotted lines indicate aligned fractures. Localities and their respectively layers are shown in the upper-right border of each picture (layout: LOG-METER; see position of samples in Fig. 4). Legend is the same for Fig. 3. Outcrops: LRO: La Rosita; MCH: Muralla China; SRP2 (Salto Río Pollux)
Calcareous-rich microfacies
Calcareous-rich microfacies comprise mostly skeletal allochems, and scarce intraclasts (Figs. 2; 3). Bioclasts are reworked and dominated by bivalves (mostly oysters), calcareous red algae, and echinoid fragments (Figs. 2A–F; 3A). Red algae is controlled by Archamphiroa jurassica Steinmann (1930), and scarce Parachaetetes sp. and “solenoporoids” (Bucur, pers. com., 2020; Fig. 2E, F). Less abundant bioclasts include reworked serpulids (Fig. 3A), bryozoans (Fig. 3G) and, rarely, benthic foraminifers (Fig. 3F) and ostracods. Non-skeletal allochem comprise intraclasts composed of bioclastic wackestone (intraclasts type 1, Fig. 3B), or formed of “cloudy” radiaxial fibrous calcite (intraclasts type 2; Fig. 3C). Bioclasts are sub-angular- to sub-rounded, usually affected by microboring, or sometimes conforming cortoids (Fig. 2B, F).

Petrography and components. A, C–D, F, K–L Calcareous-rich microfacies; B, G–J, M–O volcaniclastic-rich microfacies. A Mixed, pack–rudstone wit oysters and encrusted serpulids; note oxidation of the argillaceous-calcareous matrix (microfacies Mf-1). B Intraclasts type 1: bioclastic wackestone bearing the same fossils found discretely in the framework; note compaction, pressure solution and organic-rich matrix (microfacies Mf-4). C Intraclasts type 2: reworked clasts formed of radiaxial calcite (microfacies Mf-2, 3). D Fragment of carbonized wood (microfacies Mf-3a). E Left: calcisphere from microfacies Mf-1; right: phosphatic algal cysts (acritarch?) from organic matter-rich microfacies Mf-4om. F Several benthic foraminifers from microfacies Mf-2, 3 and 4. G Reworked and strongly bioeroded bryozoan colony, from a devitrified volcaniclastic-rich sample (microfacies Mf-4); zoecia have been filled with clay- and silica cements. H Organic matter-rich matrix from microfacies Mf-4om; note pressure solution and concentric alteration of volcanic lithics. I Similar composition observed in volcaniclastic-rich microfacies Mf-4; note difficult differentiation of lithics and matrix as well as formation of clay and chert cements, likely after devitrification. J Compositional variability of volcanic lithics. Left: intermediate clast with trachytic texture (microfacies Mf-4); right: rhyolitic clasts with micropoikilitic texture (devitrified; microfacies Mf-4c). K Common marine calcareous cements found in the mixed rocks: isopachous dogtooth around a bioclast (black arrow); porosity subsequently filled by blocky-type cement. L Pseudo-grainstone with a strong neomorphic texture; note the presence of mud peloids, not clear if primary of secondary (pseudomatrix; microfacies Mf-1). M–O close-up of volcaniclastic-rich lithofacies Mf-4 (quartzo-feldspatho-lithic sandstone); note similar composition of volcanic lithics and matrix. M–N Compaction of matrix forming pressure solution seams (white arrow); note possible devitrified-replaced relict pumice clasts (vesicular? black arrow), and formation of argillaceous cements around clasts (sericite). O Picture displaying similar composition between volcanic lithics (dotted lines) and matrix, interpreted as originally vitreous (pyroclastic?); note white mica in the central part (white arrow)
Matrix is neomorphic (microspar), but patches of mud-peloids and micritic envelopes were also observed (Fig. 3L). Intergranular porosity was filled by thin layers of isopachous and dog-toothed calcite, followed by granular, blocky, and syntaxial cements (Flügel 2010; Figs. 2C, E, F; 3K, L).
Terrigenous-rich microfacies
The terrigenous part is dominated by oligomictic, non-vesicular volcanic clasts and isolated crystals (Fig. 2C, I-K). Volcanic rock fragments are sub-angular to sub-rounded; they have an intermediate composition (quartz-trachyte) with a trachytic-porphyritic texture (potassium feldspar phenocrysts, minor plagioclase, scarce pyroxene and quartz; Fig. 3I, J). In contrast, felsic rock fragments are less frequent (rhyolite, rhyodacite < 10%), displaying a felsitic or micropoikilitic texture (Fig. 3J). Volcanic groundmass is pale brown-colored, partly devitrified or sericitized, or locally replaced to carbonates, Fe oxides-hydroxides, silica (chalcedony), or scattered opaques (Figs. 2A, D, K; 3I, J). Discrete crystals are dominated by euhedral–subhedral feldspars (mostly sanidine, minor orthoclase and plagioclase), slightly to moderately altered (Figs. 2K; 3I). Quartz is angular to sub-rounded, sometimes displaying resorption (Figs. 2J, K; 3I). Infrequent minerals include micas (mostly biotite; Figs. 2J, 3G), and rare rounded glauconite.
The terrigenous matrix is clay-silt-sized, brown- to gray-colored (Figs. 2G, H; 3G–I), sometimes stained by Fe oxides-hydroxides (Figs. 2A, B). It bears silt-sized, angular crystals and opaque minerals, and it usually displays solution seams (Figs. 2G, H; 3H). In crossed nicols, it appears isotropic or partly sericitized, hindering its differentiation from discrete lithics and reflecting a similar composition with them (Fig. 3I, M–O). Locally, some altered-compacted lithics have been converted to pseudomatrix (Fig. 2A). Cementation is minor, including silica and clay cements (Figs. 2K; 3G), and wispy-shaped chert (Fig. 3I); but also, scattered calcite or dolomite (Figs. 2C, I–K, 3J).
Microfacies
Five microfacies were defined (microfacies Mf-1 to Mf-5); they are briefly described below and summarized in Table 1. These will be referred to in the following sections, linked to their associated lithofacies and outcrops. Detailed compositional information of each thin section can be found as supplementary material (Table SM-4).
Microfacies Mf-1: It comprises sandy–gravelly, volcaniclastic–bioclastic packstone to rudstone, with grainstone patches (Fig. 2A, B). Its fabric is poorly washed and massive, but locally imbricated (bivalves). Framework is rich in bioclasts and altered trachytic rock fragments embedded in argillaceous microspar, locally associated with mud-peloids (Fig. 3L), or clayey matrix (pseudomatrix, Fig. 2A, B). Bioclasts includes mostly oysters, followed by red algae and echinoids; serpulids (some of them attached to oysters; Fig. 3A) and intraclasts type 2 are sporadic; bryozoans and calcispheres are scarce (Fig. 3E).
Microfacies Mf-2: Formed of sandy–gravelly, volcaniclastic–bioclastic float–wackestone, with packstone patches (Fig. 2D). This microfacies is analog in fabric and allochems to Mf-1, though poorer sorted, richer in big-sized bioclasts, and poorer in volcanic lithics. Additionally, its neomorphism is stronger, with coarser microspar (pseudo-grainstone), calcite veins and solution seams (“fitted” packstone; Fig. 2D). Novel fossils include scarce inoceramid fragments.
Microfacies Mf-3: It comprises sandy volcaniclastic–bioclastic grainstone, and red algae-rich grainstone (sub-type Mf-3a; Fig., 2F), with rud-, wacke-, or packstone patches (Fig. 2E, F). Its fabric is massive or locally showing aligned bioclasts. In contrast to previous microfacies, this one is poorer in volcanic clasts, and allochems are better rounded and sorted. Bioclasts comprise mostly bivalves, red algae, and subordinate echinoids plates (Fig. 2E, F). Less frequent allochems include intraclasts type 2 (Fig. 3C), and scarce bryozoans, carbonized wood (Fig. 3D), benthic foraminifers (Fig. 3F), and ostracods. Matrix is patchy (microspar, mud-peloidal; Fig. 2F), or it occurs as micritic envelopes; cementation is analog to previous microfacies (see Sect. “Calcareous-rich microfacies”), though more abundant (Fig. 3K).
Microfacies Mf-4: This microfacies is composed of gravelly-muddy, mixed bioclastic–volcaniclastic, quartzo-feldspatho-lithic to feldspatho-lithic sandstone (Figs. 2C, I, J; 3I, M–O). Microfacies Mf-4 is poorly sorted, either rich in devitrified matrix, or bearing organic matter (Mf-4om; Figs. 2G, 3E, H). Some samples also show patches of calcitic cements (granular and poikilotopic, sub-type Mf-4c; Fig. 2C, I). Its framework is mostly composed of sub-angular to sub-rounded volcanic lithics, and angular to sub-angular crystals. It displays a subtle parallel lamination (Fig., 2I, J), noticed by aligned elongated bioclasts or micas (mostly biotite, minor white mica).
Volcanic lithics are mostly intermediate, but some samples bear considerable felsic clasts (proportion intermediate:felsic = 9:1 to 6:4; Fig. 3I). Many rock fragments concentrate opaques in the core (Figs. 2G; 3H) or, when devitrified, they are replaced by chert (Fig. 3I), sericite, or calcite (pumice? Fig. 3M, N). Bioclasts comprise mostly reworked bivalves, and minor red algae and echinoids (Fig., 2C, G, I); bryozoans and intraclasts type 1 are scarce (Fig. 3B). Matrix is argillaceous-carbonaceous, and cements were already described (see “Terrigenous-rich microfacies”; Figs. 2I, K; 3G; M–O). Solution seams are common, concentrating insoluble residues and organic matter (Figs. 2G, 3H).
Microfacies Mf-5: Formed of (muddy) volcaniclastic quartzo-feldspathic sandstone with subtle parallel lamination (Fig. 2J, K). Its framework is fine- to medium-grained, dominated by angular to sub-angular crystals (mostly feldspars, followed by quartz, and minor lithics and micas). Volcanic lithics are subordinated, usually devitrified, or altered (Fig. 2K). Argillaceous matrix is scarce. Cements include calcite/dolomite, usually spotted or altering the framework (Fig., 2K); titanite was also observed. Bioclast and micas are elongated and imbricated; grains appear fractured in a preferential direction.
Interpretation of microfacies
Calcareous-rich microfacies reflect a major carbonate sedimentation incorporating reworked, older-altered volcanic rocks, or sporadic input of fresh volcaniclastics. Carbonate mud might have an allochthonous origin (mud-peloids, bioeroded clasts, absence of fecal pellets), and a minor bacterial origin (cortoids, microbial structures). Cementation depicts a phreatic marine environment and the subsequent porosity-filling during burial diagenesis (Flügel 2010). However, intraclasts type 2 may indicate erosion of aragonite-filled cavities in a coastal setting (later recrystallized to calcite; see Kendall and Tucker 1973). Coarse-grained, rud-floatstone microfacies (Mf-1, Mf-2) reflect a near-coast reworking influenced by currents (aligned bioclasts) and a rapid settling (poorly-sorted). Grainstones (Mf-3) depict a near-coast, good hydraulic sorting (Flügel 2010).
Terrigenous-rich microfacies indicate a submarine-settled, volcaniclastic sedimentation. For microfacies Mf-4, rounded lithics and abundant shell debris reflect epiclastic transport or coastal reworking. Non-vesicular volcanic rock fragments, and their core-margin differential alteration may be related to phreatomagmatism or quenching, respectively (Fisher and Schmincke 1984; Cas and Wright 1987). Allochems are analog to the ones found in the calcareous microfacies (Mf-1 to Mf-3), they were most likely swept off- (shell debris) or eroded from near-coast areas (cemented?). Given their poor textural maturity (matrix-rich) and devitrified matrix, these microfacies are interpreted as formed after remobilization of non-consolidated deposits. However, organic-rich samples (e.g., phosphatic algal cysts in Mf-4om; Fig. 3E) indicate sedimentation in a poorly oxygenated setting or during a period of high productivity (Capelli et al. 2021). Microfacies Mf-5 depicts remobilization of crystal-rich volcaniclastics, likely fractionated during the eruption or concentrated during the epiclastic transport (see Chapter 11 in Cas and Wright 1987). Its parallel lamination, and the incorporation of bioclasts is linked to current-transport. Calcareous cements are interpreted as diagenetical, while fractured crystals and titanite might reflect burial diagenesis and circulation of high-T pore fluids.
Lithofacies
Six lithofacies were defined in the study area; they are presented in decreasing grain-size order (Table 2). The main lithology is shown in upper case (e.g., R: rudstone; SG: sandstone > conglomerate, also sandy conglomerate), whereas lower case prefixes indicate composition (e.g., b: bioclastic; v: volcaniclastic). Suffixes refer to structures (e.g., p: planar bedding; m: massive). Sedimentary logs and their associated lithofacies can be found in Fig. 4.

Detailed view and correlation of sedimentary logs, including approximate stratigraphic position of the lithofacies (left size of the logs), and samples (small rhombs to the right). A La Rosita outcrop. B Muralla China outcrop. C Salto Río Pollux outcrop. See “Lithofacies” and “Sedimentary logs and facies associations”. The legend is valid for the following illustrations (Figs. 5, 6, 7). Scale is the same for all outcrops
Mixed bioclastic–volcaniclastic, sandy conglomerate (bGSm)
This lithofacies is formed of bioclastic–volcaniclastic, sandy conglomerate, arranged in tabular beds (Fig. 5). Bedding is 0.3–0.4 m-thick, with sharp contacts, usually marked by changes in gravel content (Fig. 5I). The framework of lithofacies bGSm is matrix-supported (locally clast-supported), conformed by pebble-sized clasts (diameter: max = 2.5 cm, average = 1 cm), embedded in a coarse- to very coarse-grained, sandy–muddy calcareous matrix (Fig. 5E, I, J). Exceptionally, a singular bed of lithofacies bGSm bears cobble-sized lithics (sub-facies bGSm-g; clasts-size up to 15 cm, average 2–5 cm; Fig., 5G). Gravel is formed of massive to diffusely graded, volcanic rocks and frequent bioclasts (Fig. 5E, G, J). Lithics are poorly-sorted, rounded- to sub-angular, light-colored and mono-oligomictic. Fossils are reworked, highly fragmented, and poorly packed; bioclasts are dominated by bedding-concordant, platy oyster shells (length up to 10 cm; Fig. 5E), though some beds also bear in situ and reworked colonial corals (Fig. 5J) as well as scarce carbonized wood remains.

La Rosita outcrop (LRO). A Simplified stratigraphic log; the complete one, including its lithofacies, is presented in Fig. 4A. B Panoramic view of the mixed calcareous outcrops, including the one here studied at the lower-right margin (white rectangle). C Close-up of the upper part of the outcrop, displaying a decreasing bed-thickness upwards (white arrow), and the alternation of sharp-sinuous and amalgamated contacts. D Crudely/diffusely bedded strata of the LRO section, showing decreasing bed-thickness (white arrow) and differential weathering of beds forming overhangs (close-up of the white square from B); person as scale (black arrow). E Coarse-grained lithofacies composed of gravel-sized oyster fragments and volcanic lithics (lithofacies bGSm). F Erosive contact between the mixed lithofacies of the Toqui Formation (Manto Member) and the Foitzick Volcanic Complex, a local member of the Ibáñez Formation. In G, I, J: Coarse-grained mixed lithofacies showing terrigenous material (black arrows: gravel-sized volcanic clasts) and bioclasts (white arrows). G Bioclastic-volcaniclastic, pebble-cobble-sized sandy conglomerate (lithofacies bGSm-g). H Reworked and bioeroded, big-sized gryphaeid oyster (Aetostreon sp.1) I Contact between lithofacies bGSm (mixed bioclasticvolcaniclastic, sandy conglomerate) and bSGm (mixed bioclastic–volcaniclastic, gravelly sandstone); white arrows point to reworked corals. J Close-up of lithofacies bGSm; note the preservation of colonial corals in life position (white arrows) between the gravel (black arrows). Scale bar = 1 cm. Legend for all pictures and sedimentary logs is presented in Fig. 4
The matrix of lithofacies bGSm consists of sandy–gravelly, volcaniclastic–bioclastic, packstone–rudstone, with grainstone patches (microfacies Mf-1; Fig. 2A, B).
Interpretation: Based on its mixed volcaniclastic–calcareous composition, coarse-grained and poorly sorted components, and reworked sessile and free-living epifauna, lithofacies bGSm represents deposition in a relatively high-energy coastal environment (Leithold and Bourgeois 1984; Plint 2010). Coarse-grained coastal sediments are usually linked to the reworking of fluvial mouth-bar deposits (Bourgeois and Leithold 1984; Hart and Plint 1995), though the mono-oligomictic composition of the volcanic clasts suggests erosion of a local source, as observed in some volcanic islands (Felton 2002; Felton et al. 2006). The tabular beds, poor sorting, and matrix-supported fabric, may indicate a rapid settling, likely occurring during storms (Leithold and Bourgeois 1984); the latter is also supported by the packstone and rudstone microfacies (Flügel 2010). Mixture of sub-rounded and sub-angular clasts might indicate abrasion in the beachface, during storm swells (“swash area”; Hart and Plint 1989; 1995).
Oyster-rich rudstone/coquinite (bRp)
The lithofacies bRp consists of an accumulation of cobble-sized, broken bivalve shells (40%), mostly oysters. Beds are sharp-bounded and decimeter-thick. Shells are fragmentary, platy- and wedge-shaped, arranged chaotically or concordant with bedding (Fig. 6I). They are clast-supported and embedded in a light grey-colored, fine-grained calcareous matrix.
Interpretation: This lithofacies corresponds to a basal lag formed by shell debris (Collinson et al. 2006). Based on its coarse grain-size, dense packing, and bedding-concordant bioclasts, it is interpreted as a “sedimentologic concentration” deposit, conforming proximal tempestites settled over the normal-weather wave base (Kidwell et al. 1986), and likely transported by storm-induced bottom flows (Flügel 2010).
Mixed bioclastic–volcaniclastic, gravelly sandstone (bSGm)
This lithofacies resembles the mixed conglomerate facies (bGSm), but it is finer grained. It consists of bioclastic–volcaniclastic, gravelly sandstone to granule-pebble-sized, sandy conglomerate (Fig. 5I). Strata of lithofacies bSGm are decimeter-thick, crudely bedded, amalgamated or grading from the gravel-rich ones (bGSm), though some sharp bed-boundaries are sinuous (Fig. 5C, D). Internal structures were not observed; they were likely effaced during weathering (Fig. 5D). Sandstone is medium- to very coarse-grained, whereas conglomerate is granule- to pebble-sized (clasts-size up to 1–2 cm, average < 1 cm), and usually sandy matrix-supported. Its framework is rich in volcanic lithics and fragmented oyster shells (length < 2 cm), and minor corals in some strata (Fig. 5I).
Under the microscope, the matrix of lithofacies bSGm varies between sandy, volcaniclastic–bioclastic limestone (microfacies Mf-1; Fig. 2B), and gravelly, bioclastic–volcaniclastic, quartzo-feldspatho-lithic sandstone (microfacies Mf-4c; Fig. 2C).
Interpretation: Analog to bGSm, lithofacies bSGm depicts a subtidal environment. Better sorting of clasts and oriented shell debris reflect current-influenced hydraulic sorting. Sinuous-topped, amalgamated beds depict periodic erosion–deposition pulses (Collinson et al. 2006), and subsequently wave- or current reworking (Walker and Plint 1992; Clifton 2006); whereas sharp contacts might represent reactivation surfaces. These tabular beds could represent the formation of gravelly–sandy low relief bedforms linked to alongshore currents (similar to Fig. 14 in Hart and Plint 1995; also “bedload-sheets” sensu Carling 1999; or “carbonate sandbodies” sensu Wright and Burchette 1996). These bedforms are typically developed on the shoreface (Hart and Plint 1995).
Rippled, bioclastic (volcaniclastic) limestone (bLr)
The lithofacies bLr is composed of gravelly–sandy, bioclastic limestone, with a minor volcaniclastic content. It is arranged in tabular layers with sinuous bed boundaries, usually amalgamated (Fig. 6C–E). Based on the grain-size and scale of ripples, two subtypes were defined: (i) bLr-m: “mega-rippled” (sensu Swift et al. 1983), or coarse-grained-rippled (sensu Leckie 1988), gravelly, bioclastic limestone (Fig. 6D, E); and (ii) bLr-o: oscillation-rippled, sandy, bioclastic limestone (Fig. 6F, G).

Muralla China outcrop (MCH). A Overview of the outcrop, displaying the contact relationship with the Ibáñez Fm. and the position of the “Muralla China sill”; black square: MCH outcrop. B Simplified stratigraphic log (complete profile in Fig. 4B). C Close-up of the MCH outcrop from A (Jacob’s staff as scale within white circle, 1.5 m-length); red circles with capital letters are linked to the pictures D-N. D megarippled bioclastic limestone (lithofacies bLr-m); note decimeter wave-length and amalgamated contacts. Black arrows: sharp erosive contacts (sinuous). E Gravelly–sandy bioclastic limestone (bSGm) with amalgamated contacts (dotted line) from the lowermost part of the outcrop; note sinuous contacts (black arrows) and scattered bioclasts. F Several parallel beds conformed of oscillation-rippled, sandy bioclastic limestone (lithofacies bLr-o); note some amalgamated contacts and parallel-arranged tafoni (white and black arrow, respectively). G Close-up of the previous, in a different part of the outcrop; rippled surfaces are marked with black arrows. H Reworked left valves of oysters (Aetostreon sp. and A. sp.2; black arrows), from a tempestite in the uppermost part of the outcrop. I Oyster coquinites from the northern part of the outcrop (lithofacies bRp). J An ex-situ cobble including a cluster of Aetostreon sp.2, likely fallen from the uppermost part of the outcrop (see H). K Reworked small pectinid (Entolium? sp.). L Carbonized fossil word forming a thin-bedded coal bed. M Reworked, inequivalve, big-sized oyster. N Reworked, bioeroded, and articulated big-sized oyster (Aetostreon sp.1; also displayed in Fig. 9A–B). Scale bar (white) = 1 cm
The mega-rippled subtype (bLr-m) consists of sandy–gravelly float–rudstone, bearing pebble- to cobble-sized bioclasts and volcanic lithics (1–2 cm-length, max = 9 cm-length in broken-disarticulated oyster shells). The framework is massively arranged, but, locally, it displays a sub-parallel arrangement of shells (Fig. 6E). Limestone is sharply bedded, arranged in decimeter to meter-thick bedsets with sinuous contacts (Fig. 6D). Internally, the sub-type bLr-m displays symmetric large-sized ripples (wavelength ≈ 50–90 cm; height ≈ 15–20 cm; Fig. 6D). On the other hand, the oscillation-rippled subtype (bLr-o) is formed by mixed, gravelly–sandy calcarenites (grain size 1–5 mm, max: 1.7 cm), displaying smaller and regular symmetric ripples, and subordinate asymmetric ripples (wavelength ≈ 20–30 cm; height ≈ 10 cm; Fig. 6F, G).
The matrix of lithofacies bLr-m consists of sandy–gravelly, volcaniclastic–bioclastic float–wackestone with packstone patches (microfacies Mf-2; Fig. 2D), whereas sub-type bLr-o is composed of sandy, bioclastic rudstone–grainstone, and red algae-rich grainstone (microfacies Mf-3, Mf-3a; Fig. 2E, F).
Interpretation: This lithofacies depicts a shallow marine environment emplaced above the fair-weather-wave base (Burchette and Wright 1992; Flügel 2010). Mega-ripples (bLr-m) have been regarded as the equivalent of hummocky cross-stratification for coarse-grained sediments, which are typically linked to storm-surge deposits (Leckie 1988; Cummings et al. 2009). Based on the previous, and on their poorly sorted microfacies (Mf-2), they are interpreted here as calcareous proximal tempestites (analog to “Facies A” of Pérez-López and Pérez-Valera 2012). On the other hand, given its grain size, better sorted microfacies (Mf-3), and symmetrical rippled-beds, lithofacies bLr-o reflects an oscillatory flow typical of fair-weather wave action, and minor influence of near-coast unidirectional currents (Collinson et al. 2006; Plint 2010).
Parallel-bedded, bioclastic–volcaniclastic, muddy sandstone (bSp)
Lithofacies bSp crops out as sandy, bioclastic mudstone to calcareous muddy sandstone; it is dark grey-colored, and it expels a fetid odor when crushed (Fig. 7C, E). Its bedding is arranged in decimeter-thick layers with diffuse, flat-amalgamated contacts (10–40 cm-thick; Fig. 7A-C, E, H). Beds are commonly friable; however, some better cemented strata serve as guide-beds, these have sharp margins with a rippled top or base (sub-facies bSp-c; Fig. 7B–D, H). Bedding is internally massive, but some beds display a coarse-tail grading of bioclasts (i.e., muddier tops, or diffuse amalgamated ripples (Fig. 7D). Shell debris are abundant, regularly to well-sorted, and highly fragmented (ca. 20–30% bioclasts, 2–3 mm-length; Fig. 7E); they comprise mostly bed-concordant bivalves, and some echinoids fragments. Other complete macrofossils are scattered, including reworked oysters and belemnites (Fig. 7F, G, I, J).

Salto Río Pollux outcrop (SRP2). A Simplified sedimentary log. B–C Overall view of the outcrop displaying the inclined bedding, amalgamated beds, and intrusive contacts with the Muralla China sill (red dashed line). Light-blue arrows: calcareous beds; grey arrow: tuffaceous sandstone (lithofacies tSp-b); white arrow: decreasing bed-thickness upsection. D Overhang of carbonate-cemented bed of lithofacies bSp-c, exposed between brittle bedsets of lithofacies bSp; note the sinuous internal, amalgamated surfaces (hammer as scale). This layer corresponds to the lowermost light-blue arrow in B–C. E Close-up of lithofacies bSp (bioclastic, muddy sandstone); note its dark-colour and matrix-rich composition as well as abundant comminuted shell debris (pencil as scale). F Reworked (convex-up) and bioeroded specimen of Aetostreon sp.2 within lithofacies bSp. G Complete, well-preserved specimen of Belemnopsis sp. H Close-up of C, displaying the middle position of the tuffaceous strata (lithofacies tSp-b); note diffuse cross-bedded? beds of calcareous sandstone (bSp-c) above. I Aetostreon sp.2 sampled from lithofacies tSp-b; the fossil stills bear sediments. J Gryphaeid oysters (Aetostreon sp.2) in life position conforming a small cluster. Scale bar (white) = 1 cm
Microscopically, lithofacies bSp consists of gravelly–muddy, mixed bioclastic–volcaniclastic sandstone (microfacies Mf-4, Mf-5 subordinate).
Interpretation: Based on its amalgamated tabular beds, parallel lamination (microfacies Mf-4, Mf-5), sandy framework, bimodal bioclasts (fragmented-complete), and grading, this lithofacies reflects waning flows with an intermediate character between proximal (i.e., coarse-grained, bioclast-dominated; similar to “Facies B” of Pérez-López and Pérez-Valera 2012) and distal tempestites (i.e., interbedded with mud; Flügel 2010). The latter are usually difficult to differentiate from turbidites (e.g., “turbidite-like tempestites” sensu Myrow 1992; 2005), however, turbidites are finer grained (mud to fine-sand sensu Shanmugam 1997, 2002, 2016). Its volcaniclastic composition reflects remobilization of non-consolidated deposits to relatively quiet, soft-bottom subtidal settings.
Laminated, tuffaceous muddy sandstone (tSp-b)
This lithofacies is formed of bioclasts-bearing, medium-to-coarse-grained, muddy sandstone (Fig. 7C, H, I). Sandstone is friable, light grey-colored, slightly calcareous, and partly altered to phyllosilicates. Bedding is arranged in centimeter-thick layers with parallel bedding (Fig. 7C, H), and its framework is rich in white-colored, sand-sized rounded clasts, interpreted as altered pumice (Fig. 7I). Subordinately, it bears sand-sized micas and glauconite. Its fossil content includes reworked left valves of gryphaeid oysters (Fig. 7I).
Interpretation: Based on its composition and alteration, this lithofacies is interpreted as a submarine-settled, tuffaceous sandstone (remobilized tuffaceous deposits sensu McPhie et al. 1993). Given its grain size, lamination, and fossil content, it may reflect sedimentation by currents.
Sedimentary logs and facies associations
This section describes three sedimentary logs (Fig. 1D) as well as their facies associations and paleoenvironmental interpretation. Outcrops have mixed composition varying from calcareous-rich (LRO, MCH) to volcaniclastic-rich lithofacies (SPR2); they are described from north to south. The detailed description of each sedimentary log and their beds, compiled in the field, has been provided as supplementary material (Tables SM-1 to SM-3).
La Rosita Section (LRO)—gravelly, mixed inner ramp
This locality comprises at least three grey-colored, wedge-shaped small outcrops, exposed in a ca. 1 km radius, about 8 km to the south of Coyhaique (Figs. 1D; 5B–D; 4A). The central outcrop, here described (LRO), is better exposed and consists of a 17-m-thick succession slightly inclined to the SSW (strike/dip = 105/13; Figs. 5A–D). The outcrop displays several overhangs with diffuse contacts, after amalgamation and differential weathering of the strata (Fig. 5D). These rocks include mixed, volcaniclastic–bioclastic conglomerate and sandstone (lithofacies bGSm; bSGm; Fig. 5E, G, H–J), and subordinate bioclastic–volcaniclastic sandstone (bSp, Fig. 5I).
At the base, mixed rocks overlie meter-sized, volcanic boulders of the Foitzick Volcanic Complex with an erosive unconformity (De la Cruz et al. 1994, 2003; Fig. 5F). In addition, an angular unconformity, and local peperitic contact have been reported from adjacent, correlative outcrops (Bell et al. 1994; Suárez et al. 1996; Suárez and De la Cruz 1994a, b). The top of the LRO section is not exposed (Fig. 5B–D).
In the LRO section, the mixed succession is arranged in decimeter-thick, tabular beds, commonly amalgamated (avrg. 0.3–0.4 m, max: 0.8–1.0 m, probably fused beds; Fig. 5C–D). Upsection, sediments are better sorted and their bedding is clearer (Fig. 5C). Coarsest, poorly sorted gravelly beds are usually thicker and more common in the lowermost part of the outcrop (bGSm; bGSm-g, Fig. 5E, G, J), changing to a gravelly–sandy alternation in the central part (bGSm; bSGm; Fig. 5C, D). The last third of the outcrop displays a thinning- and fining-upwards trend, dominated by gravelly sandstone with diffuse- and sharp, sinuous contacts (lithofacies bSGm; bSp; 0.3–0.5 m-thick; Fig. 5C); some beds are richer in volcaniclastics (microfacies Mf-4c, Table 1).
Macrofossils are reworked and fragmented, dominated by reworked oysters (rarely big-sized specimens of Aetostreon sp.1; Fig. 5H), but colonial corals were also found in the lowermost beds, some of them in life position (Bell et al. 1994; Townsend 1998; and here, see Fig. 5J); belemnites are scarce and broken. Articulated- and encrusting oysters in life position have also been reported from this outcrop (“Exogyra” sp. sensu Bell et al. 1994; Townsend 1998), but they were not observed during this work.
Interpretation: The La Rosita outcrop depicts a relatively shallow marine environment, placed above the fair-weather wave-base. Its sub-environments comprise a deepening-upwards succession from a storm-influenced, gravelly upper shoreface towards a sandy lower shoreface (Hart and Plint 1995). These are here interpreted to represent a mixed volcaniclastic-carbonate inner ramp (sensu Burchette and Wright 1992; Fig. 14.3 in Flügel 2010). The fragmented fossil content, including sessile and free-living epifauna, reflects the development of local communities, though not forming reefs (“ahermatypic corals” sensu De la Cruz et al. 2003), and rather grew as discrete small crusts over the coastal gravel deposits.
These gravelly deposits represent the marine erosion over a rocky shoreline (coastal cliff), eroding an older volcanic terrain (Suárez et al. 1996). Based on the inferred “onlap” contact of these rocks above the volcanic complex (illustrated in Fig. 5 in Suarez and De La Cruz 1994a; also in Fig. 3.10 in Townsend 1998; Cover Photo in De la Cruz et al. 2003; and Fig. 5B here), also reported as locally peperitic (Bell et al. 1994; Suarez and De La Cruz 1994a, b), these rocks might reflect transgressive deposits settled over a volcanic flank (De la Cruz et al. 1994; Bell et al. 1994; Suárez et al. 1996; Suárez and De la Cruz 1994a, b). In particular, the gravelly facies (bGSm) may represent transgressive conglomerates (Suárez et al. 1996).
Muralla China Section (MCH)—wave- and storm-dominated inner-ramp
This section is located 2.8 km to the south-southeast of section LRO, near the northern end of a sub-horizontal sill, locally known as “Muralla China” (Figs. 1D; 4B; 6B–C). The succession comprises a wedge-shaped, steep outcrop formed of ca. 34 m of gravelly–sandy, bioclastic limestone dipping to the southeast (strike/dip = 060/20; Fig. 6A). The limestone conformably overlies pale grey-green-colored, diffusely bedded, volcanic rocks of the Foitzick Volcanic Complex (sensu Bell et al. 1994; Suárez and De la Cruz 1994a, b, see Fig. 6A). Gravel-sized volcanic clasts found within the calcareous beds suggest erosion over this basal unit. The top of the MCH succession is not exposed (Fig. 6A).
In the MCH outcrop, the lower and uppermost beds are coarser grained, conformed of volcaniclastic–bioclastic, mixed gravelly–sandstone (lithofacies bSGm; Fig. 6E), and mega-rippled bioclastic limestone (lithofacies bLr-m; Fig. 6D, F, G). The central part of the outcrop is relatively homogeneous and consists of well-sorted, wave-rippled bioclastic limestone (bLr-o; Fig. 6C, F, G). Bedding is tabular to wedge-shaped, displaying a subtle, fining- to coarsening-upwards trend (Fig. 6B). Bedding planes are commonly amalgamated, sinuous, and only recognized by tafoni superimposed on ripples (Fig. 6C, F). Sharp boundaries are subordinate, these are interpreted as erosive or reactivation surfaces (Fig. 6C).
The fossil content is fragmentary, though reworked, but articulated big-sized oysters (Gryphaeidae indet; Aetostreon sp.1.; Fig. 6M, N) and other small pectinids (Entolium? sp.; Fig. 6K), as well as bulb-shaped fish teeth (Pycnodontidae indet.), were found scattered in the middle part of the section (Fig. 6A, C). Near the top, complete, small- and medium-sized, oyster left-valves assigned to Aetostreon sp. and A. sp.2 (Fig. 6H) are abundant in mixed, gravelly floatstone beds. The latter were also found as small clusters, but detached from its original bed (ex situ; Fig. 6J). Exceptionally, some lenses of carbonized wood were observed (Fig. 6L) and, at the northern end of the outcrop, a lenticular bed of oyster-rich rudstone/coquinite was found (bRp; Fig. 6I).
Interpretation: Based on wave ripples (Walker and Plint 1992), mostly comminuted shells, and packstone–grainstone microfacies, these sediments suffered wave-reworking above the fair-weather wave-base, i.e., in the inner-ramp (Burchette and Wright 1992; Flügel 2010). This mixed inner ramp was affected by periodical storm-surges, as shown by mega-rippled limestone, coarse-grained coquinites, and layers with a bimodal distribution of bioclasts interpreted as tempestites (Kidwell et al. 1986; Flügel 2010; Fig. 6H). Storm-currents were strong enough to remobilize and rapidly bury big-sized, reclining oysters (Aetostreon spp.). These energetic events were periodic as shown by amalgamated layers and pinched-out bed-sets, enclosed by erosive surfaces (Fig. 6C).
This outcrop is described here for the first time. Regarding composition, thickness, and inferred age (Sect. “Stratigraphy and correlations”), the Muralla China limestone corresponds to one of the most remarkable Lower Cretaceous calcareous outcrop reported from the Aysén-Río Mayo Basin.
Salto Río Pollux Section (SRP2)—mixed, distal mid-ramp
This outcrop is located about 1 km to the south of the Muralla China Section (MCH), at the northern hand of Route 7, just before reaching the Pollux River (Fig. 1D). It is composed of ca. 9 m of dark-colored, bioclastic muddy sandstone dipping towards the southeast (strike/dip = 041/10; Figs. 4C; 7A, B). Its base and top are not exposed; however, the outcrop is capped by the Muralla China sill (see 1.2; Fig. 7A-C) and, about 100 m to the east, this outcrop is separated from the black mudstones of the Katterfeld Formation by an inferred fault (Fig. 1D).
The Salto Río Pollux outcrop comprises bioclastic–volcaniclastic, muddy sandstones (lithofacies bSp), alternated with calcareous sandy beds (bSp-c; Fig. 7A). The former (bSp) are more abundant in the lower half of the outcrop, exposed as poorly defined, amalgamated beds (Fig. 7C, H). Some of them are rich in organic matter (microfacies Mf-4om; Fig. 7E). In contrast, calcareous beds (bSp-c), stand out as weathering-resistant guide beds (Fig. 7B–D). Bed-boundaries between these both lithofacies are sharp and sinuous (Fig. 7C, D). The middle point of the outcrop is marked by a 0.3 m-thick, fine interbedding of fossil-bearing, mixed sandstone (bSp) and tuffaceous sandstone (lithofacies tSp-b; Fig. 7C, H, I).
Bioclasts are abundant, dominated by bedding-concordant shell debris (mostly oysters; Fig. 7E). Complete macrofossils include scattered belemnite rostra (Belemnopsis sp.), and left valves of Aetostreon sp.2 (“Aetostreon sp. nov.” Rubilar 2000; Fig. 7F, I, J here), the latter also found conforming small clusters in life position (Fig. 7J).
Interpretation: Based on the matrix-rich, mixed sandstone (microfacies Mf-4), its bimodal fossil distribution (comminuted-complete), the amalgamation of its beds, and its sedimentary structures, these deposits reflect “event sedimentation” or “eventites” (Flügel 2010). They are interpreted as mixed calcareous–volcaniclastic tempestites settled in a mixed, distal mid-ramp (Wright and Burchette 1996; Flügel 2010), i.e., between the fair-weather- and the storm wave-base (Burchette and Wright 1992; Ramalho et al. 2013).
These tempestites show grain size differences between gravelly-grained shelly fragments and sandy-sized volcaniclastics, indicating a turbulent hydraulic sorting (Kidwell and Bosence 1991; Immenhauser 2009; Figs. 3I; 7E). The abundant comminuted shelly fragments were most likely swept off from the breaker- or swash area and were offshore-transported during storm-surges (Flügel 2010). As shown from the muddy matrix and well-preserved complete fossils, storm currents interrupted the relatively quiet background sedimentation, rapidly burying (or mildly reworking) the small, thin-shelled, recliner oysters in life position (Aetostreon sp.2; Fig. 7F, I–J) as well as other corporal fossils (e.g., belemnites; Fig. 7G). These small-sized Aetostreon sp.2 likely proliferated in a water–sediment interface affected by sporadic events of eutrophication, inferred from its organic matter content (Nori and Lathuilière 2003).
The high terrigenous content of the Salto Río Pollux beds reflects a major volcaniclastic supply (Table 1; Figs. 2G–K; 3M–O). Given the interbedding with tuffaceous beds (lithofacies tSp-b; Fig. 7C, H), and the similar composition of volcanic lithics and sedimentary matrix (Microfacies Mf-4; Fig. 3I, M–O), a remobilization of non-consolidated volcaniclastic deposits is inferred (syneruptive?, McPhie et al. 1993; Schneider 2000). Previously, these volcanic deposits were classified as “volcanic coquinas” by Katz (1961). This author inferred a poorly-oxygenated, marine environment for these rocks, linked to a coeval, subaerial- and submarine volcanic eruptions. The former is supported by the fetid-odor and organic matter content of these rocks (microfacies Mf-4; O’Brien and Slatt 1990; Ulmer-Scholle et al. 2014), and it could be linked to settling in a restricted environment or during a period of high biological productivity (Capelli et al. 2021). Regarding the volcanism, there is not enough evidence to support a subaqueous source.
Systematic paleontology
Oysters and red algae are the most abundant fossils found in the outcrops analyzed here. This section presents a detailed description of newly found fragments of the calcareous red alga Archamphiroa jurassica, identified here for the first time in the Lower Cretaceous of Chile (see “Calcareous-rich microfacies”; Fig. 2E, F; 8A–I). In addition, some oysters of the Aetostreon genus are also described (Fig. 9).
Archamphiroa jurassica
Class Rhodophyceae Rabenhorst 1863
Order ?Corallinales Silva and Johansen 1986
Genus Archamphiroa Steinmann 1930; emend. Bucur et al. 2009
Archamphiroa jurassica Steinmann 1930 (Pl. 1, Figs. 1–9]); emend. Bucur et al. 2009 (neotype, Figs. 2–3).
Type material: The neotype was designated by Bucur et al. (2009) in algal wackestone of the Cotidiano Formation (see Fig. 1A, C). According to these authors, a neotype was required given the loss of the original material, in which no holotype had been defined (see Steinmann 1930).
Type locality: Puesto Cotidiano (44°50’S; 71°39’W), at the isthmus connecting the La Plata and Fontana lakes in the Chubut province of Argentina (Fig. 1C). Originally defined by Steinmann (1930) at the Arroyo Negro, tributary of the Malargüe River in the Neuquén province of Argentina (ca. 35°S).
Material: Dozens of thallus fragments from the La Rosita (LRO, five thin sections; Fig. 4A) and Salto Río Pollux outcrops (SRP2, four thin sections; Fig. 4B); hundreds of thallus fragments from the Muralla China beds (MCH, six thin sections; Fig. 4C).
Description: Because of their small size, fragmentary preservation, strong cementation, and similar color to that of the calcareous matrix-cement, thalli of A. jurassica are difficult to identify in hand samples. The following characteristics were used here for the assignation in thin section (based on Steinmann 1930; Bucur et al. 2009).
External: Thallus fragments are singular or frequently bifurcated, with rounded margins (Fig. 8A, C, E, F–I). Their cross-sections are circular, or ellipsoidal when obliquely cut (Fig. 8E, G, I); longitudinal views are ellipsoidal or sub-rectangular (Fig. 8A–C, H). When bifurcated, thalli are lambda-shaped, branches appear parallel-arranged, showing no hinge-like structure or constriction at the point of bifurcation (i.e., non-geniculate; Fig. 8F–H). Given their size and “dotted”, internal cell arrangement, thalli fragments might be confused with echinoids, though red algae display a clear fan-like extinction (Fig. 8H, I).

Red algae identified in the Toqui Formation (Manto Member). A–I Archamphiroa jurassica Steinmann (1930) A Longitudinal view displaying the internal hypothallium–perithallium contrast; note the alternation of dark–light arched layers in the hypothallium (apical part to the right). B Red algae-rich grainstone (microfacies Mf-3a), including several fragments of A. jurassica; note the bioerosion of bioclasts and dissolution-refill of the hypothallium. C Similar to A, longitudinal view of A. jurassica displaying the alternated layering (dark-coloured arches convex to the left). D Reworked, bioeroded, and micritized red algae fragments, possible source of the calcareous sediments (microfacies Mf-3a); note compaction (concave-convex contacts). E Club-shaped cavities in the peripheral zone of a reworked alga (cross-section), interpreted as bioerosion (white arrow). F Longitudinal view of A. jurassica displaying its non-geniculate ramification. G Cross-section of A. jurassica displaying the ramification (bifurcation) and hypothallium-perithallium contrast. H Longitudinal view displaying non-geniculate ramification, and the fan-shaped cell-arrangement in the hypothallium. I Circular cross-sections of thalli from A. jurassica are easy to confuse with echinoid spines; however, the red alga shows a fan-like extinction. J Reworked specimen of Parachaetetes sp. K Reworked specimen of “Solenopora” sp. Photographs from C, E, J, K courtesy of Prof. Dr. Ioan Bucur. Scale: black = 0.5 mm, white = 1 mm
Internal: Thalli of A. jurassica are two-layered, with a central hypothallium covered by an outer perithallium (Fig. 8A, C, F–I). The hypothallium is multilayered coaxial, conformed of an alternation of dark- and pale-colored arched cellular layers, convexly-oriented towards the apical part (Fig. 8A, C, F). Internally, the arches comprise elongated filaments, which diverge in an acute fan-shaped fashion (Fig. 8F, H). Filaments of the perithallium are oriented perpendicular to the thallus margin (Fig. 8A, E–G, I). In cross-section, the hypothallium appears darker or with a structureless cell arrangement, contrasting with the radial pattern observed in the perithallium (Fig. 8E, G, I).
In order to confirm peripheral cellular fusion, regarded as a diagnostic feature of this taxon, samples were observed using SEM, but without successful results. This may be linked to diagenetical alteration or incorrect preparation of the samples (see SEM-images as supplementary material). Nevertheless, the assignation was confirmed by the Prof. Dr. Bucur (pers. com., 2020), with whom an algae-rich sample was shared.

Aetostreon spp. from the area south of Coyhaique. A–L Muralla China outcrop (MCH); M–R: Salto Río Pollux outcrop (SRP2). A–B (Sample MCH-14.8/1) Aetostreon sp.1, left valve, lateral view (A = external, B = internal); C–D: (Sample MCH-33/R/8) Aetostreon sp., external lateral- (C) and dorsal views (D). E–F (Sample MCH-33/R/2) Aetostreon sp., left valve, external lateral- (E) and posterior views (F); G–I: (Sample MCH-33/16) Aetostreon sp., left valve, external lateral- (G), posterior- (H), and dorsal views (I). J–R: Aetostreon sp.2. J–K: (Sample MCH-33/12) Aetostreon sp.2, external lateral- (J) and posterior views (K); L: (Sample MCH/R/5) Aetostreon sp.2, internal mold; M–N: (Sample SRP2-3.2/9) Aetostreon sp.2, left valve, lateral external- (M) and posterior views (N); O–P: (Sample SRP2-2.7/6) Aetostreon sp.2, left valve, lateral external- (O) and posterior views (P); Q-R: (Sample SRP2-2.7/2) Aetostreon sp.2, left valve, lateral external- (Q) and internal views (R). Abbreviations: am adductor muscle scar (short dashed line); be bioerosion (long dashed line); oc opisthogyrate coiling; pk posterior keel; pr paradontal recess; uc umbonal cavity
Dimensions: The main size parameters for A. jurassica are summarized in Table 3 (n = 128, single measures are presented in Table SM-5 as supplementary material):
Remarks:
Reproductive structures are absent. However, many thallus fragments show occasional circular, club-shaped, and irregular cavities as well as small, marginal vermicular borings, caused by macro- and microborers (Fig. 8A–B, E).
Based on its cylindrical thallus shape and non-geniculate bifurcation, a fruticose to arborescent growth form is inferred for Archamphiroa jurassica (sensu classification of Woelkerling et al. 1993; see Fig. 10A, A’).

(Modified from Rivas et al. 2021). Given the different composition and possible tectonic contacts between outcrops, which may reflect different timing of sedimentation (see “Discussion” in text), they are presented separated between calcareous-rich (LRO, MCH) and volcaniclastic-rich (SRP2). A Mixed, calcareous-volcaniclastic ramp model, during sedimentation of the LRO- and MCH beds. A’ Cross-section displaying the shoreline-offshore arrangement of the facies associations, and the hypothetically, original habitat (communities) of the taxa here described as a fossil assemblage. B Mixed, calcareous–volcaniclastic ramp model, during sedimentation of the SRP2 beds. B’ Cross-section displaying the shoreline-offshore arrangement of the facies associations. I-R inner ramp, M-R mid-ramp, O-R outer ramp, ad non-genicular algal debris, as the result of near-coast wave and storm-reworking, bioerosion, and grazing; they might resemble the current “maerl-type” deposits; ap algal pavements/meadows developed in the inner ramp, likely conforming non-geniculate rhodoliths; cc coastal carbonates; source of the eroded and crushed material, offshore-transported towards the deeper facies (SPR2); dt distal tempestites, formed by the offshore transport (mid-ramp) of shell-debris and sand-sized volcaniclastics during major storm-surges; ol small-sized, gryphaeid oysters in life position (isolated or in clusters; see Figs. 6, 7); rc rocky coast (source of gravel and encrusting organisms); ro reworked gryphaeid oysters; rv paralic/coastal resedimented volcaniclastics, interpreted as the main source of siliciclastic material in mixed lithofacies; sv syneruptive volcaniclastic input, eruption-fed (syneruptive) or after costal reworking of non-consolidated tuffaceous deposits. Scales are approximate, based on the current position of the outcrops (fossils not to scale)
Model for depositional environments and paleoecology of the Toqui Formation from the Coyhaique area
Stratigraphic range: An early Callovian age was inferred for the beds bearing Archamphiroa jurassica in Central Argentina by Steinmann (1930), based on its association with Upper Jurassic brachiopods (“Rhynchonella acuticosta”) and corals (Thamnasteria sp.). In Southern Argentina, a late Tithonian age was suggested for the Cotidiano Formation by Bucur et al. (2009), unit bearing the A. jurassica neotype, type specimen defined by the same authors. The latter was based on the presence of Steinmanella (Macrotrigonia sp.) and Megatrigonia cf. fontanaensis, and on the close similarity of this fauna with the one found at Arroyo Pedregoso, which has been ammonite-dated to late Tithonian by Olivero (1982; based on Berriasella sp. and Micracanthoceras ruedai).
Outside southern Argentina, Archamphiroa jurassica Steinmann (1930) is now described from Coyhaique, Chile. This red alga is abundant in the mixed-calcareous outcrops here presented (see Sects. “La Rosita Section (LRO) - gravelly, mixed inner ramp”–“Muralla China Section (MCH) - wave and storm-dominated inner-ramp” above). The age of these outcrops is restricted to the late Berriasian/early Valanginian -Hauterivian? (see “Age of the deposits”). Therefore, this extends the range of A. jurassica to the Early Cretaceous, in particular, between the Tithonian-Valanginian (Hauterivian?).
Aetostreon spp.
Superfamily Ostreoidea Rafinesque 1815
Family Gryphaeidae Vialov 1936
Subfamily Exogyrinae Vialov 1936
Tribe Nanogyrini Malchus 1990
Genus Aetostreon Bayle 1878
Type species: Gryphaea latissima Lamarck 1801, p. 399.
Diagnosis (modified from Stenzel 1971; Cooper 1995; Toscano and Lazo 2020): Medium- to large-sized, thick-shelled, inequivalve. Opisthogyrate umbo, gyrostreoid or exogyroid ligament area. Left valve: convex and deep; right valve: flat or slightly concave. Left valve thickened in the crest zone of umbonal ridge, displaying a keel in the posterior third. Keel is rounded to acute, commonly surmounted by knobs. Shallow groove parallel to the keel separates a slightly more convex posterior flank. No ornamentation except for growth lines or occasionally radial wrinkles. Internally, adductor muscle scar usually subtruncate antero-dorsally, but sometimes rounded; paradontal apparatus well developed; chomata lacking.
Description: Oysters assigned to Aetostreon in this study display a marked morphological variation, interpreted here as evidence of the occurrence of at least two different species. Morphs are discernable based on their size and general shape of the valves, but specific relevant taxonomic information is unfortunately not preserved. Shells of Aetostreon sp.1 are big-sized, thick, with a subtrigonal outline and a keel more pronounced in the dorsal third (Figs. 5H; 9A-B); internally, lack of an umbonal cavity, and the rounded adductor muscle scar is dorsally truncated (Fig. 9B). Overall, Aetostreon sp.1 resembles A. latissimum from the northern Neuquén Basin in Argentina (e.g., Lazo 2007; Rubilar and Lazo 2009; Aguirre-Urreta et al. 2011; Toscano and Lazo 2020). The lack of key diagnostic traits (i.e., ligament area, umbo), presently demands the use of an open nomenclature for them.
Specimens of Aetostreon sp.2 are smaller, and thinner than A. sp.1, their left valves are globular with a subovate outline, and the umbo is opisthogyrate, with a small- to big-sized attachment area (Fig. 9J-R), and well-preserved growth lines (Fig. 9M, O, P). The keel is more pronounced in the umbonal area, changing to a slightly more convex posterior flank in the rest of the valve (Fig. 9L-R). Internally, Aetostreon sp.2 displays a moderate umbonal cavity (Fig. 9L, R), a narrow, linear paradontal recess, a subcircular adductor muscle scar, and the lack of chomata (Fig. 9L, R); these features are very similar to “Aetostreon sp. nov.” of Rubilar (2000). A third type of Aetostreon displays intermediate size between both Aetostreon sp.1 and A. sp.2, with left valves elongated in a ventral or ventro-posterior direction (Fig. 9C, E), a subovate- to subtriangular outline (Fig. 9C, E, G), and a keel in the posterior third, more pronounced near the opisthogyrate umbo (Fig. 9D–E, G). Given their poor preservation and sediment-coverage, it is not clear whether these oysters represent juveniles of Aetostreon sp.1, bigger morphotypes of Aetostreon sp.2, or a third different species. Therefore, they are presented conservatively as Aetostreon sp. (Fig. 9C–I).
Stratigraphic range: Globally, Aetostreon have been reported from the Oxfordian-Albian, but mainly from the Berriasian-Aptian range (Cooper 1995; Toscano and Lazo 2020, and references therein). In the Neuquén Basin of Argentina, several species of Aetostreon have been described in early-mid Berriasian (A. subsinuatum), late Berriasian-early Valanginian (A. latissimum; Toscano and Lazo 2020), late Valanginian (A. pilmatuegrosum; Rubilar and Lazo 2009), and late Valanginian-early Hauterivian units (Aetostreon sp.; Lazo 2007).
Discussion
Paleogeography
In the Aysén-Río Mayo Basin, several mixed- and calcareous outcrops have been identified (e.g.,Ploszkiewicz and Ramos 1977; Ramos 1978; Olivero 1983, 1987; Scasso 1987; Covacevich et al. 1994; Bell et al. 1994; Suárez et al. 1996; Townsend 1998; Ramos and Lazo 2006; Suárez and De la Cruz 1994a, b, among many others). However, an unequivocal correlation of these deposits has been hindered, due to their discrete and discontinuous exposures, rapid lateral and vertical facies changes (Scasso 1989; Scasso and Kiessling 2002), and diachronism (Suárez et al. 2005, 2010a). The latter is linked to a marine incursion over an irregular topography, affected by extensional tectonics and active volcanism (Ramos 1981; Scasso 1987; 1989; Folguera and Iannizzotto 2004; Suárez et al. 2009, 2010a). In addition, during the Cenozoic, strong strike-slip tectonic disruption as well as Neogene glacial events have affected the Coyhaique area (Scalabrino 2009). This is evident in the study area, where the outcrops are disconnected, incomplete, and located at different elevations (Figs. 1C, D; 5B, 6A, 7B).
Nevertheless, based on composition, common fossil assemblages, inferred sedimentary environments, and contact relationship with the volcanic units, the outcrops La Rosita (LRO) and Muralla China (MCH) appear to be laterally correlative, and part of the same carbonate-rich, mixed inner ramp setting (Figs. 4A, B; 10A–A’). Volcaniclastic-rich strata from the uppermost LRO outcrop show a sporadic clastic input, whereas thicker bedded limestones of the MCH outcrop reflects a relatively stable carbonate sedimentation, either located in a more distant setting from the coastal volcaniclastic source, or deposited during a phase of quiescent terrigenous supply (Nelson 1988; James 1997; Dorobek, 2008; Fig. 10A). Both outcrops are carbonate-rich and assigned here to the Manto Member of the Toqui Formation (sensu Rivas et al. 2021).
As shown in Sects. “La Rosita Section (LRO) - gravelly, mixed inner ramp”–“Muralla China Section (MCH) - wave- and storm-dominated inner-ramp”, this ramp was developed during the marine transgression above a volcanic terrain (Fig. 10A, B), interpreted by several authors as an insular volcanic flank (e.g., Bell et al. 1994; De la Cruz et al. 1994; Suárez et al. 1996; Townsend 1998; De la Cruz et al. 2003; Suárez et al. 2023). The latter may explain the preservation of gravel-rich facies, common in volcanic coasts affected by rapid shoreline translation (Felton 2002; Ramalho et al. 2013).
The relationship between the previous outcrops (LRO, MCH) and the Salto Río Pollux (SRP2) section is unclear (Fig. 4B, C), given their contrasting lithologies, structural attitude, and inferred tectonic contacts. Although they share the same fossil content (but in different proportions, see Table 1), which may support a lateral correlation, the SRP2 succession bear intraclasts type 1 (reworked bioclastic limestone; Fig. 3B), and its composition reflects a deeper setting and a major volcaniclastic supply. Alternatively, based on the volcaniclastic-rich facies of the SRP2 outcrop, more compatible with the San Antonio Member of the Toqui Formation (sensu Rivas et al. 2021), it might represent a vertical transition in the facies, with sediments deposited in a (younger) period controlled by a terrigenous and probably syneruptive input (tuffaceous beds of lithofacies tSp-b; Fig. 10B–B’). An increased clastic supply may also be related to enhanced run-off, triggered by the change from dry to humid (and hotter?) conditions (Capelli et al. 2021).
Paleoenvironmental interpretation
“Cool-water” carbonate setting
Following the “cool-water” criteria of James (1997), and based on paleolatitude (non-tropical), platform morphology (ramp-type), and mode of life of benthic organisms (heterozoan association), a non-tropical, “temperate” or “cool-water” (sensu lato, Schlager 2003) setting is inferred for the study area, i.e., an open marine environment with water temperatures of < 18°–20 °C (“temperate-type” sensu Simone and Carannante 1988; James 1997; Schlager 2005). However, since factors other than temperature may strongly affect carbonate ecosystems and their grain associations (e.g., terrigenous input, trophic conditions, water chemistry, CO2 saturation, among others, Kindler and Wilson 2010; Westphal et al. 2010; Michel et al. 2018), a “cool water” setting is used here in a broad sense (see contrasting examples in “Regional comparison”). Criteria supporting a cool-water setting for the rocks studied here are explained below.
As shown in “Age of the deposits”, the mixed rocks studied here appear to be restricted to the Lower Cretaceous. For that period, paleotectonic reconstructions of South America place the study area to south of 45°–50°S (e.g., Leinfelder et al. 2002; Scasso and Kiessling 2002; Seton et al. 2012; Matthews et al. 2016), i.e., in a non-tropical, high paleo-latitude (Scasso and Kiessling 2002).
Facies, sedimentary structures and reworked bioclasts depict a shallow-marine environment for the mixed outcrops of the Toqui Formation (refer to Sects. “Sedimentary logs and facies associations”). Wave- and storm-related structures reflect a relatively high-energy and a hydrodynamic control (Figs. 5C, 6C-G; 7D, E) regarded as typical for open shelves and ramp-type carbonate platforms lacking physical barriers (i.e., reefs, sand shoals; Burchette and Wright 1992; Wright and Burchette 1996; James 1997; Pedley and Carannante 2006; Flügel 2010).
Tropical carbonates usually accumulate in low latitudes (between ca. 30°N and S of the equator, Schlager 2003), in waters with temperatures higher than 20°–22 °C (Nelson 1988; James 1997; Schlager 2003). They are characterized by photo-autotrophic organisms with a dominant aragonitic mineralogy (e.g., green and red algae, hermatypic corals, and photosymbiotic interactions), and a biotic-abiotic precipitation of carbonate mud, cements, and non-skeletal allochems (Lees and Buller 1972; Nelson 1988; Schlager 2003; Kindler and Wilson 2010 and references therein). In contrast, limestones documented here are poor in carbonate mud and non-skeletal allochems (only sporadic intraclasts), and the benthic fauna is dominated by calcareous red algae and calcitic-shelled heterotrophic organisms (see next Sect. “Paleoecology”), conforming a heterozoan association (sensu James 1997; Clarke 2009; Kindler and Wilson 2010).
Paleoecology
The fossil assemblage studied here is mostly composed of sessile organisms, including encrusters (red algae; serpulids; bryozoans; ahermatypic corals, oysters; Figs. 2; 5E, H, J), soft-bottoms recliners (gryphaeid oysters; Figs. 6J, M, N; 7F, J), vagile epifauna (echinoids, pectinids; Fig. 6K), and nektonic animals (fishes, belemnites; Fig. 7G). Most of these organisms are stenohaline and support an euhaline and shallow, open marine environment (e.g., Wray 1977; Scholle and Ulmer-Scholle 2003; Stenzel 1971; Toscano et al. 2018; Cawley et al. 2021). However, some of these organisms may also have an euryhaline physiology (e.g., bryozoans, oysters, red algae, serpulids sensu Scholle and Ulmer-Scholle 2003), thus, local brackish areas or zones with a variable salinity may also be feasible (Townsend 1998). This has been commonly reported from some volcanic coastlines with an alluvial influence (Ramalho et al. 2013).
Red algae are the most remarkable photo-autotrophic organisms found in these rocks of the Toqui Formation (Fig. 8); they might represent the main source of carbonate (Wray 1977; Flügel 2010; abraded-bioeroded algae in Fig. 8B, D). Algal content is reworked and found as broken branches or rounded bioclasts, abundant in wave-influenced sediments (inner ramp; lithofacies bLr). An original shallow-water setting in the photic zone is inferred for them (Michel et al. 2018), encrusting submerged rocky blocks (“solenoporoids”), or conforming semi-protected?, low-relief meadows or “pavements” in coastal areas (see Fig. 5 in Henrich et al. 1995; Fig. 10A, A’ here). The latter, rich in ramified non-geniculate algae (A. jurassica), is considered as wave-reworked and offshore-transported (Fig. 10A, A’). Red algae-rich grainstones like the ones described resemble the current “maerl-type” deposits, conformed by broken branches of non-geniculate, unattached rhodoliths, typically found today in high-latitude, shallow and cool-water settings of the North East Atlantic (Henrich et al. 1995; Ehrhold et al. 2021).
Ahermatypic corals flourished during the incipient transgression, encrusting subtidal gravel in a shallow-water setting; they disappeared upsection likely due to sediment stress (Risk and Edinger 2011; see Fig. 10A, A’). Echinoid plates and spines are rich in wave- and storm-reworked sediments. An epifaunal or semi-infaunal habit in the sandy sediments, grazing on the algal meadows is commonly recognized for them (Nebelsick and Bassi 2000; Kroh 2003; see Fig. 10A, A’).
In the Aysén-Río Mayo Basin, oysters are mostly represented by exogyrids of the genre Aetostreon (Fig. 9), regarded as a homeomorph of Gryphaea (Stenzel 1971). Gryphaeids could live attached or free-lying (MacKerrow, 1981) in subtidal settings with soft-bottom and euhaline quiet water (Stenzel 1971; LaBarbera 1981). Analogous to Gryphaea, Aetostreon is also depicted as a soft-bottom recliner (Lazo 2007) living in shallow, low-energy inner-shelf settings with sandy–muddy, soft-bottoms (Cooper 1995; Lazo et al. 2005; Rubilar and Lazo 2009); in intertidal shelly mudstones (Cooper 1995); or forming biostromes in micritic mid-ramp settings (Rivas et al. 2021). In this study, big-sized articulated Aetostreon sp.1 from the LRO and MCH outcrops are found reworked in sandy limestones (lithofacies bLr), reflecting a local provenance (Figs. 5H, 6M-N, 9A–B). It is unclear if the oyster coquinites (lithofacies bRp) from the MCH outcrop may correspond to the same species (Fig. 6I), while in the LRO outcrop, Townsend (1998) reported life-positioned oysters encrusted in the basal volcanic blocks, but without providing a taxonomical assignation.
Smaller oysters from the MCH and SRP2 outcrops were tentatively classified as different species (Aetostreon sp. and A. sp.2; Fig. 9), though subtle morphological differences between specimens may represent morphotypes (Rubilar 2000). For example, the uppermost beds of the MCH outcrop bear a mixture of medium-sized (Aetostreon sp.; Fig. 9C–I) and small-sized oysters (Aetostreon sp.2; Fig. 9J–L); both appear as reworked and disarticulated valves, embedded in sandy limestone (Fig. 6H, J). In contrast, in the SRP2 outcrop, only small specimens were observed (Aetostreon sp.2 = “Aetostreon sp. nov.” Rubilar 2000, ex “Gryphaea balli” sensu Cecioni and Charrier 1974; Fig. 9M-R), in life position and reworked, embedded in shelly, muddy sandstone (lithofacies bSp; Fig. 7F, I–J).
In Gryphaea, their relatively heavy shells are unlikely to be distantly transported, serving as a good paleoecological indicator (Bayer et al. 1985). In addition, wider morphotypes may reflect an ecophenotypic adaptation to firmer substrates (e.g., sand-rich); this is also supported by the presence of bioeroded and reworked shells, more exposed to erosional events (Bayer et al. 1985). Therefore, in the MCH outcrop, small- and mid-sized Aetostreon spp. were likely transported from low-energy, muddy–sandy shallow-marine settings (see paragraph above), or they were adapted to bioclastic sandy bottoms of the carbonate inner ramp (Fig. 10A, A’). In the SPR2 beds, dominant small-sized Aetostreon sp.2 thrived in mud-rich soft-bottoms (LaBarbera 1981), with tendency to eutrophication (rich in organic matter; Nori and Lathuilière 2003), interpreted here as part of the mid-ramp (Fig. 10B, B’).
Based on the previous, the fossil assemblage described here depicts an autochthonous-para-autochthonous assemblage in the inner-ramp (outcrops LRO, MCH; Fig. 10A, A’), but an autochthonous-allochthonous in the mid-ramp (outcrop SPR2; Fig. 10B, B’).
Paleoclimate
The calcareous sedimentation in the Aysén-Río Mayo Basin was strongly diachronic, likely ranging from the Tithonian–Valanginian, or even Hauterivian (see Sect. “Stratigraphy and correlations”). Paleoclimate models for the Jurassic-Cretaceous transitions are usually contrasting. Some authors infer an equable warm climate, with a lower temperature gradient and absence of polar ice caps (Hay 2008; Föllmi 2012), while others depict episodic cooling or “cool-snaps” (Price 1999; Kessels et al. 2006; Donnadieu et al. 2011), or even seasonal growth of polar ice (Price 1999, 2009; Price et al. 2000; McArthur et al. 2007; Mutterlose et al. 2009; Hay and Floegel 2012; Tennant et al. 2017 among many others). Globally, the most remarkable J-K cool-spans are depicted as occurring during the late Tithonian, the mid-Valanginian, and across the Aptian-Albian boundary (Price 1999), though they have being called into question in more recent studies (Föllmi 2012; Jenkyns et al. 2012). This paradox is also extended to southern South America, where some models infer warm (> 25 °C), sea-surface temperatures in the Southern Ocean during the J-K transition (ca. 52°S; Jenkyns et al. 2012; Vickers et al. 2019); while other propose episodic cool intervals in the late Tithonian–earliest Berriasian, Valanginian, earliest Hauterivian, late Barremian, Aptian, and earliest Albian (Salazar and Stinnesbeck 2015; Brysch 2018).
In the Aysén-Río Mayo Basin (Fig. 1B), the paleoclimatic evidence for the existence of cold snaps is inconclusive. Overall, warm and humid conditions have been inferred for the deposition of the Coyhaique Group (Tithonian–Aptian), but only based on the presence of limestone (Skarmeta 1976; Bell et al. 1994, 1996; Townsend 1998). However, as presented in “Paleoecology”, mixed rocks of the Toqui Formation exposed south of Coyhaique, and assigned to the late Berriasian/early Valanginian-Hauterivian? (see “Age of the deposits”; “Stratigraphy and correlations”), display typical features of “cool water” carbonates (see ““Cool-water” carbonate setting”).
Two events of global cooling are known from this period and are supported by evidence from Patagonia (Brysch 2018). The first one, across the Berriasian-Valanginian boundary, is marked by a glacioeustatic, long-term sea-level fall (Pucéat et al. 2003; Haq 2014). The second, in the Valanginian (Weissert and Erba 2004; Cavalheiro et al. 2021), corresponds to a positive carbon isotope excursion (“Weissert Event”; e.g., van de Schootbrugge et al. 2000; Meissner et al. 2015; Price et al. 2018), linked to a high rate of organic carbon burial (Erba et al. 2004; Price et al. 2018). Alternatively, these “episodes of environmental change” (sensu Föllmi 2012), among others during the Early Cretaceous, may represent short-termed humid periods alternated with long-termed arid greenhouse conditions (Föllmi 2012). For example, to the north, in the Neuquén basin (ca. 37°S), a shift to more humid conditions have been inferred during the early Berriasian–latest early Valanginian, prior to the onset of the Weissert Event (Capelli et al. 2021). There, humid conditions would have led to an enhanced runoff, triggering a shift from calcareous-rich to clastic-rich sedimentation (Capelli et al. 2021) as observed here (see ““Cool-water” carbonate setting”).
However, given the several unresolved problems regarding the global J-K paleoclimate models (Hay 2017), and the ambiguous, broad age range inferred for these outcrops of the Toqui Formation, more analytical data are required, to incorporate them in the global paleoclimatic scheme.
Regional comparison
The Toqui formation has been correlated with the upper Tithonian Cotidiano Formation, and with the Tithonian? – Hauterivian Tres Lagunas Formation from Argentina (45°S; see Sect. “Stratigraphy and correlations”). Archamphiroa jurassica has been reported from both the Cotidiano Fm. and from the outcrops presented here (Toqui Fm.), though both units differ in their carbonate depositional settings and inferred ages (see “Age of the deposits”). The Cotidiano Formation consists of biohermal limestone (“coral-stromatoporoid patch reefs”; Ramos 1978), analog to the “Photozoan Association” of James (1997), likely settled in a tropical–subtropical environment (T > 22 °C sensu Ramos 1976; 1978), or in a warm-water ocean (T > 18 °C sensu Kiessling et al. 1999; Leinfelder et al. 2002; Leinfelder et al. 2005). This contrast may be explained by the marked temporal gap between both deposits (Tithonian versus upper Berriasian-Hauterivian?), and potential deposition under differing paleoenvironmental conditions.
A similar problem arises when comparing the present outcrops of the Toqui Formation with those from the Tres Lagunas Formation. Even though both units display a mixed carbonate-clastic composition and partly coeval ages, their calcareous facies and depositional environments are clearly different. At its type locality (Laguna Salada), the Tres Lagunas Formation includes conglomerates, limestones (coquinites and coral-bioherms), tuffaceous beds, and sandstone–mudstone alternations (some calcareous or bioclastic); these rocks are discretely exposed showing abrupt vertical and lateral facies changes (Scasso 1987; 1989; Scasso and Kiessling 2002). Carbonate facies of the Tres Lagunas Fm. have been interpreted as coral patch-reefs and associated fore-reef deposits (Olivero 1983; Scasso 1987; 1989), likely developed in a warm-water setting during the Berriasian-Valanginian (Scasso 1989; Scasso and Kiessling 2002). These rocks depict a rimmed carbonate shelf (bioherms), contrasting with the wave- and storm-influenced, open-marine ramp deposits presented here (see Sects. 4, and 6.1).
The cause behind these opposing depositional environments is not clear. Paleolatitude and temporality do not seem to be the controlling factors, and other variables must be considered; for example, their position within the basin (see paleo-reconstructions in Scasso 1989; Folguera and Iannizzotto 2004), but also coast morphology and hydraulic settings (rimmed shelf vs ramp profile), coeval volcanism (Scasso and Kiessling 2002), and terrigenous input or trophic conditions (e.g., coastal upwelling; Michel et al. 2018). The latter could have played a role in reef growth (Scasso and Kiessling 2002), as observed repeatedly in volcanic islands (Ramalho et al. 2013). Several other factors may favor the development of photozoan versus heterozoan benthic communities (e.g., temperature, salinity, water depth, CO2 concentration, etc.; Nelson 1988; Pomar et al. 2004; Kindler and Wilson 2010; and references therein); their role in the study area is still unclear and needs to be addressed in future studies.
Conclusions
Three sections assigned to the Lower Cretaceous Toqui Formation, the lowermost member of the Coyhaique Group, are here investigated from an area south of Coyhaique in southern Chile (45°40’S). These rocks overlie the volcanic Ibáñez Formation, regarded to represent the early Valanginian in the study area, and their top is not exposed. In the study area, the sedimentological and microfacial analysis of the Toqui formation reveals a mixed calcareous–volcaniclastic composition, rich in reworked bioclasts conforming a heterozoan-type fossil assemblage. Mixed lithofacies and associated sedimentary structures reflect the development of a wave- and storm-influenced mixed ramp. Based on their lithologies, two outcrops were assigned to the Manto Member (calcareous-rich LRO, MCH sections) and one to the San Antonio Member of the Toqui Formation (volcaniclastic-rich SRP2 section). The calcareous sedimentation is depicted as part of a retrograding coast above an active volcanic terrain with sporadic activity.
Bioclastic, mixed calcareous rocks of the Toqui Formation conform a “heterozoan” association (heterotrophic organisms + red algae), bearing gryphaeid oysters (Aetostreon spp.), non-geniculate calcareous red algae, and echinoids as major constituents. Red algae are controlled by Archamphiroa jurassica, identified here for the first time in the Lower Cretaceous of Chile and previously reported only from the Tithonian of Argentina.
Based on the inferred high paleolatitude of these rocks (south of 45°–50°S), their heterozoan fossil assemblage, and depicted depositional setting (open-marine, ramp-type), a “cool-water” non-tropical setting is inferred for the Aysén-Río Mayo Basin in the Coyhaique area during the late Berriasian/early Valanginian—Hauterivian? This contrasts with the depositional environments inferred for correlative reefal rocks in Argentina (Tres Lagunas Formation), which reflect “warm-water” settings. In the Aysén-Río Mayo Basin, the influence of sea-water key physical variables in the carbonate sedimentation, as well as the position of the carbonate platforms within the basin, and their interaction with the volcanism are still unclear.
Data availability statement
The data that support the findings of this study are available as supplementary material of this article. This additional information includes: i) description of each sedimentary log and their beds, compiled in the field (Table SM-1 to SM-3); ii) description and modal composition of each thin section (Table SM-4); measurement of exemplars of Archamphiroa jurassica under the microscope (Table SM-5).
References
Aguirre-Urreta MB, Ramos VA (1981) Estratigrafía y Paleontología de la Alta Cuenca del río Roble, Cordillera Patagónica—Provincia de Santa Cruz. In: VIII Congr. Geol. Argentino, Actas, Buenos Aires, Argentina, III, 101–133.
Aguirre-Urreta MB, Suárez M, Bruce Z, De la Cruz R, Ramos VA (2000) Bioestratigrafía y amonoideos de la Formación Katterfeld, (Cretácico Inferior) en Puerto Ibáñez, XI región, Chile. In: IX Congr. Geol. Chileno, Actas, Puerto Varas, Chile, 2, 183–187
Aguirre-Urreta MB, Lazo DG, Griffin M, Vennari V, Parras AM, Cataldo, C., Garberoglio, R. and Luci, L. (2011) Megainvertebrados del Cretácico y su importancia bioestratigráfica. In: Geología y Recursos Naturales de la Provincia del Neuquén, Asociación Geológica Argentina, Buenos Aires, Argentina, pp 465–488
Aguirre-Urreta MB (2002) Invertebrados del Cretácico Inferior. In: Geología y Recursos Naturales de Santa Cruz (Ed. MJ Haller). Relatorio del XV Congreso Geológico Argentino, El Calafate, Argentina, pp 439–459
Artemieva IM (2023) Back-arc basins: a global view from geophysical synthesis and analysis. Earth Sci Rev 236:104242
Bayer U, Johnson ALA, Brannan J (1985) Ecological patterns in Middle jurassic Gryphaea: The relationship between form and environment. In: Sedimentary and Evolutionary Cycles (Ed. U. Bayer and A. Seilacher), Springer, Berlin, Heidelberg, 1, 436–463
Bayle K (1878) Fossiles principaux des terrains: Explication carte geologique France. 158 pp.
Bell CM, Suárez M (1997) The Lower Cretaceous Apeleg Formation of the Aisen basin, Southern Chile. Tidal sandbar deposits of an epicontinental sea. Andean Geology 24:203–225
Bell CM, Townsend MJ, Suárez M, De la Cruz R (1994) The depositional environments of the Lower Cretaceous Coyhaique Group, Aysén Basin, southern Chile (45°-46°S). In: VII Congr. Geol. Chileno, Actas. Concepción, Chile, pp 402–403
Bell CM, De la Cruz R, Suárez M, Townsend MJ (1996) The evolution of the Aysen Basin, an early Cretaceous epicontinental interior seaway in the southernmost South America. Géodynamique andine: résumés étendus. ORSTOM, Paris, pp 289–292
Blair TC, McPherson JG (1999) Grain-size and textural classification of coarse sedimentary particles. J Sed Res 69:6–19
Bourgeois J, Leithold EL (1984) Wave-worked conglomerates: Depositional processes and criteria for recognition. In: Sedimentology of gravels and conglomerates (Ed. E.H. Koster, R.J. Steel, and Canadian Society of Petroleum Geologists), Canadian Soc. Petrol. Geol., Calgary, Canada, 331–343.
Brysch S (2018) Changes in climate and palaeoenvironment during the Late Jurassic–Early Cretaceous in southern South America and western Antarctica. PhD Thesis, Ruprecht-Karls-Universität Heidelberg
Bucur II, Kiessling W, Scasso RA (2009) Re-description and neotypification of Archamphiroa jurassica Steinmann 1930, a calcareous red alga from the Jurassic of Argentina. J Paleontol 83:962–968
Burchette TP, Wright VP (1992) Carbonate ramp depositional systems. Sed Geol 79:3–57
Capelli IA, Scasso RA, Spangenberg JE, Kietzmann DA, Cravero F, Duperron M, Adatte T (2021) Mineralogy and geochemistry of deeply-buried marine sediments of the Vaca Muerta-Quintuco system in the Neuquén Basin (Chacay Melehue section), Argentina: Paleoclimatic and paleoenvironmental implications for the global Tithonian-Valanginian reconstructions. J S Am Earth Sci 107:103103
Carling P (1999) Subaqueous Gravel Dunes. SEPM JSR. https://doi.org/10.1306/D4268A2C-2B26-11D7-8648000102C1865D
Cas RAF, Wright JV (1987) Volcanic successions: modern and ancient. a geological approach to processes, products and successions. Chapman & Hall, London, p 528
Cavalheiro L, Wagner T, Steinig S, Bottini C, Dummann W, Esegbue O, Gambacorta G, Giraldo-Gómez V, Farnsworth A, Flögel S, Hofmann P, Lunt DJ, Rethemeyer J, Torricelli S, Erba E (2021) Impact of global cooling on Early Cretaceous high pCO2 world during the Weissert Event. Nat Commun 12:5411
Cawley JJ, Marramà G, Carnevale G, Villafaña JA, López-Romero FA, Kriwet J (2021) Rise and fall of pycnodontiformes: diversity, competition and extinction of a successful fish clade. Ecol Evol 11:1769–1796
Cecioni, G. and Charrier, R. (1974) Relaciones entre la Cuenca Patagónica, la Cuenca Andina y el Canal de Mozambique. Ameghiniana, XI, 1–38.
Clarke J (2009) Carbonates, Cool Water. In: Gornitz V (ed) Encyclopedia of paleoclimatology and ancient environments. Springer, Netherlands, Dordrecht, pp 138–143
Clifton, H.E. (2006) A reexamination of facies models for clastic shorelines. In: Facies Models Revisited (Ed. H.W. Posamentier and R.G. Walker), SEPM, Tulsa, OK, 293–338.
Cohen KM, Finney SC, Gibbard PL, Fan J-X (2013) The ICS international chronostratigraphic chart. Episodes 36:199–204
Collinson JD, Mountney NP, Thompson DB (2006) Sedimentary structures, 3rd edn. Terra, Harpenden, Hertfordshire, p 292
Cooper MR (1995) Exogyrid oysters (Bivalvia: Gryphaeoidea) from the Cretaceous of southeast Africa. Durban Museum Novitates 20:1–48
Covacevich V, De la Cruz R, Suárez M (1994) Primer hallazgo de fauna del Berriasiano inferior (Neocomiano) en la Formación Ibáñez, región XI, Aisén. In: VII Congr. Geol. Chileno, Actas, Concepcion, Chile, I, 425–429.
Cummings DI, Dumas S, Dalrymple RW (2009) Fine-grained versus coarse-grained wave ripples generated experimentally under large-scale oscillatory flow. J Sed Res 79:83–93
De la Cruz R, Suárez M, Demant A (1994) Facies volcánicas del Mesozoico de Aisén (sector noreste), 44°-47° lat. S., Chile (Formaciones Ibáñez y Divisadero). In: VII Congr. Geol. Chileno, Actas, Concepción, Chile, I, 27–31.
De la Cruz R, Suárez M, Covacevich V, Quiroz D (1996) Estratigrafía de la zona de Palena y Futaleufú (43°15’-43°45’ latitud S), X Región, Chile. In: XIII Congr. Geol. Argentino y III Congr. Explor. Hidroc., Actas, Buenos Aires, Argentina, I, pp 417–424
De la Cruz R, Suárez M, Belmar M, Quiroz D, Bell M (2003) Geología del área Coihaique-Balmaceda, Región Aisén del General Carlos Ibáñez del Campo. SERNAGEOMIN, Santiago, Chile, 40 pp.
De la Cruz R, Welkner D, Suárez M, Quiroz D (2004) Geología del Área Oriental de las Hojas Cochrane y Villa O’Higgins, Región Aisén del General Carlos Ibáñez del Campo. SERNAGEOMIN, Santiago, Chile, 57 pp.
Demant A, Suárez M, De la Cruz R, Bruguier O (2010) Early Cretaceous surtseyan volcanoes of the Baño Nuevo Volcanic Complex (Aysén Basin, Eastern Central Patagonian Cordillera, Chile). Geol Acta 8:207–219
Donnadieu Y, Dromart G, Goddéris Y, Pucéat E, Brigaud B, Dera G, Dumas C, Olivier N (2011) A mechanism for brief glacial episodes in the Mesozoic greenhouse. Paleoceanography, 26:10
Echaurren A, Folguera A, Gianni G, Orts D, Tassara A, Encinas A, Giménez M, Valencia V (2016) Tectonic evolution of the North Patagonian Andes (41°–44° S) through recognition of syntectonic strata. Tectonophysics 677–678:99–114
Ehrhold A, Jouet G, Le Roy P, Jorry SJ, Grall J, Reixach T, Lambert C, Gregoire G, Goslin J, Roubi A, Penaud A, Vidal M, Siano R (2021) Fossil maerl beds as coastal indicators of late Holocene palaeo-environmental evolution in the Bay of Brest (Western France). Palaeogeogr Palaeoclimatol Palaeoecol 577:110525
Erba E, Bartolini A, Larson RL (2004) Valanginian Weissert oceanic anoxic event. Geol. 32, 149un
Felton EA (2002) Sedimentology of rocky shorelines: 1. A review of the problem, with analytical methods, and insights gained from the Hulopoe Gravel and the modern rocky shoreline of Lanai. Hawaii Sedimentary Geology 152:221–245
Fisher RV (1961) Proposed Classification of Volcaniclastic Sediments and Rocks. Geol Soc America Bull 72:1409
Fisher RV (1966) Rocks composed of volcanic fragments and their classification. Earth-Sci Rev 1:287–298
Fisher RV, Schmincke H-U (1984) Pyroclastic rocks. Springer, Berlin, p 472
Flügel E (2010) Microfacies of carbonate rocks: analysis, interpretation and application, 2nd edn. Springer, Heidelberg; New York, 984 pp
Folguera A, Iannizzotto NF (2004) The lagos La Plata and Fontana fold-and-thrust belt: long-lived orogenesis at the edge of western Patagonia. J S Am Earth Sci 16:541–566
Föllmi KB (2012) Early Cretaceous life, climate and anoxia. Cretac Res 35:230–257
Garzanti E (2019) Petrographic classification of sand and sandstone. Earth Sci Rev 192:545–563
Gianni GM, Navarrete C, Echaurren A, Díaz M, Butler KL, Horton BK, Encinas A, Folguera A (2019) Northward propagation of Andean genesis: insights from early cretaceous synorogenic deposits in the Aysén-Río Mayo basin. Gondwana Res 77:238–259
Gonzalez-Bonorino G, Suárez M (1995) Paleoambientes sedimentarios de la Formacion Apeleg, Cretacico Inferior de la Cuenca de Aisen, Region XI. Chile Andean Geol 22:115–126
Haller MJ, Lapido OR (1980) El Mesozoico de la Cordillera Patagónica Central. Rev Asoc Geol Argentina, XXXV, 230–247.
Haller MJ, Lapido OR, Lizuaín A, Page R (1981) El mar tithono-neocomiano en la evolución de la Cordillera Norpatagónica. In: Cuencas Sedimentarias del Jurásico y Cretácico de América del Sur, Com. Sudam. Jura. Cretac., 1, 221–237.
Haller MJ, Lapido OR (1982) The Jurassic-Cretaceous volcanism in the septentrional Patagonian Andes. Earth Sci Rev 18:395–410
Haq BU (2014) Cretaceous Eustasy Revisited. Global Planet Sci 113:44–58
Hart BS, Plint AG (1989) Gravelly shoreface deposits: a comparison of modern and ancient facies sequences. Sedimentology 36:551–557
Hart BS, Plint AG (1995) Gravelly shoreface and beachface deposits. In: Plint AG (ed) Sedimentary facies analysis. Blackwell, Oxford, UK, pp 75–99
Hay WW (2008) Evolving ideas about the Cretaceous climate and ocean circulation. Cretac Res 29:725–753
Hay WW (2017) Toward understanding cretaceous climate—an updated review. Sci China Earth Sci 60:5–19
Hay WW, Floegel S (2012) New thoughts about the Cretaceous climate and oceans. Earth Sci Rev 115:262–272
Hechem J, Figari E, Homovc J (1993) Secuencias deposicionales en el Neocomiano del lago Fontana, Chubut, Argentina. In: XII Congr. Geol. Argentino y II Congr. Explor. Hidroc., Actas, Mendoza, Argentina, II, 119–123.
Henrich R, Freiwald A, Betzler C, Bader B, Schäfer P, Samtleben C, Brachert TC, Wehrmann A, Zankl H, Kühlmann DHH (1995) Controls on modern carbonate sedimentation on warm-temperate to arctic coasts, shelves and seamounts in the Northern Hemisphere: implications for fossil counterparts. Facies 32:71–108
Hervé F, Calderón M, Faúndez V (2008) The metamorphic complexes of the Patagonian and Fuegian Andes. Geol Acta 6(1):43–45
Iannizzotto NF, Folguera A, Leal PR, Iaffa D (2004) Control tectónico de las secuencias volcaniclásticas neocomianas y paleogeografía en la zona del Lago La Plata (45°S). Sector interno de la faja plegada y corrida de los lagos La Plata y Fontana. Rev Asoc Geol Argentina 59:655–670
Immenhauser A (2009) Estimating palaeo-water depth from the physical rock record. Earth Sci Rev 96:107–139
Ippolitov AP, Desai B, Arkadiev VV (2015) First find of Parabelemnopsis, the alien belemnite from the southern hemisphere, in the upper Berriasian of Central Crimea. In: The International Scientific Conference on the Jurassic/Cretaceous boundary. Proceedings volume, Samara, Russia, 52–59.
James NP (1997) The cool-water carbonate depositional realm. In: Cool-Water Carbonates (Ed. N.P. James and J. Clarke), SEPM, Tulsa, OK, 1–20.
Jenkyns HC, Schouten-Huibers L, Schouten S, Sinninghe Damsté JS (2012) Warm middle jurassic-early cretaceous high-latitude sea-surface temperatures from the Southern Ocean. Clim past 8:215–226
Katz H (1961) Sobre la ocurrencia de Cretáceo Superior marino en Coyhaique, Provincia de Aisén. Anales De La Facultad De Ciencias Físicas y Matemáticas, Universidad De Chile, Santiago, Chile 21:113–131
Kendall AC, Tucker ME (1973) Radiaxial fibrous calcite: a replacement after acicular carbonate. Sedimentology 20:365–389
Kesjár D, Fözy I, Price G, Condon D, Salazar C, Pálfy J (2017) Integrated Lower Cretaceous stratigraphy from the Aisén Basin, Patagonia, Chile. In: 10th International Symposium on the Cretaceous - Abstracts (Ed. B. Sames), Berichte der Geologischen Bundesanstalt, Vienna, Austria, 120, 142.
Kessels K, Mutterlose J, Michalzik D (2006) Early Cretaceous (Valanginian—Hauterivian) calcareous nannofossils and isotopes of the northern hemisphere: proxies for the understanding of Cretaceous climate. Lethaia 39:157–172
Kidwell SM, Bosence DWJ (1991) Taphonomy and Time-Averaging of Marine Shelly Faunas. In: Allison PA, Briggs DEG (eds) Taphonomy: releasing the data locked in the fossil record. Plenum Press, New York, pp 115–193
Kidwell SM, Fursich FT, Aigner T (1986) Conceptual framework for the analysis and classification of fossil concentrations. Palaios 1:228
Kiessling W, Flügel E, Golonka J (1999) Paleoreef maps: evaluation of a comprehensive database of Phanerozoic reefs. AAPG Bull 83:1552–1587
Kindler P, Wilson MEJ (2010) Carbonate grain associations: their use and environmental significance, a brief review. In: Carbonate systems during the Oligocene-Miocene climatic transition (Ed. M. Mutti, W.E. Piller, and C. Betzler), Blackwell Pub, Hoboken, NJ, 35–48.
Kroh, A. (2003) Palaeobiology and biogeography of a Danian echinoid fauna of Lower Austria. In: Echinoderm research 2001: proceedings of the sixth European Conference on Echinoderm Research (Ed. J.-P. Féral and B. David), A.A. Balkema Publishers, Banyuls-sur-Mer, France, 69–75.
LaBarbera M (1981) The Ecology of Mesozoic Gryphaea, Exogyra, and Ilymatogyra (Bivalvia: Mollusca) in a Modern Ocean. Paleobiology 7:510–526
Lamarck JB (1801) Système des animaux sans vertèbres. Deterville, Paris, p 432
Lazo DG (2007) Early Cretaceous bivalves of the Neuquén Basin, west-central Argentina: notes on taxonomy, palaeobiogeography and palaeoecology. Geol J 42:127–142
Lazo DG, Cichowolski M, Rodríguez DL, Aguirre-Urreta MB (2005) Lithofacies, palaeoecology and palaeoenvironments of the Agrio Formation, Lower Cretaceous of the Neuquén Basin, Argentina. J Geol Soc London Spec Publ 252:295–315
Leckie D (1988) Wave-formed, coarse-grained ripples and their relationship to hummocky cross-stratification. J Sed Res. https://doi.org/10.1306/212F8E04-2B24-11D7-8648000102C1865D
Lees A, Buller AT (1972) Modern temperate-water and warm-water shelf carbonate sediments contrasted. Mar Geol 13:M67–M73
Leinfelder RR, Schlagintweit F, Werner W, Ebli O, Nose M, Schmid DU, Hughes GW (2005) Significance of stromatoporoids in Jurassic reefs and carbonate platforms—concepts and implications. Facies 51:288–326
Leinfelder RR, Schmid DU, Nose M, Werner W (2002) Jurassic reef patterns—The expression of a changing globe. In: Phanerozoic Reef Patterns (Ed. Wolfgang. Kiessling, Erik. Flügel, and Jan. Golonka), SEPM (Society for Sedimentary Geology), 465–520.
Leithold EL, Bourgeois J (1984) Characteristics of coarse-grained sequences deposited in nearshore, wave-dominated environments-examples from the Miocene of south-west Oregon. Sedimentology 31:749–775
Malchus, N. (1990) Revision der Kreide-Austern (Bivalvia: Pteriomorphia) Ägyptens (Biostratigraphie, Systematik). Berliner Geowissenschaftliche Abhandlungen, Reihe A, 1–231.
Masiuk V, Nakayama C (1978) Sedimentitas marinas mesozoicas del lago Fontana: Su importancia. In: VII Congr. Geol. Argentino, Actas, Neuquén, Argentina, II, 361–378.
Matthews KJ, Maloney KT, Zahirovic S, Williams SE, Seton M, Müller RD (2016) Global plate boundary evolution and kinematics since the late Paleozoic. Global Planet Change 146:226–250
MacKerrow WS (ed) (1981) The Ecology of Fossils: an illustrated guide. the MIT Press, Cambridge, Massachusetts, 384 pp.
McArthur JM, Janssen NMM, Reboulet S, Leng MJ, Thirlwall MF, van de Schootbrugge B (2007) Palaeotemperatures, polar ice-volume, and isotope stratigraphy (Mg/Ca, δ18O, δ13C, 87Sr/86Sr): the early cretaceous (Berriasian, Valanginian, Hauterivian). Palaeogeogr Palaeoclimatol Palaeoecol 248:391–430
McPhie J, Doyle M, Allen RL (1993) Volcanic textures: a guide to the interpretation of textures in volcanic rocks. Centre Ore Depos. Explor. Stud. Univ, Tasmania, Hobart, 198 pp
Meissner P, Mutterlose J, Bodin S (2015) Latitudinal temperature trends in the northern hemisphere during the Early Cretaceous (Valanginian–Hauterivian). Palaeogeogr Palaeoclimatol Palaeoecol 424:17–39
Michel J, Borgomano J, Reijmer JJG (2018) Heterozoan carbonates: When, where and why? A synthesis on parameters controlling carbonate production and occurrences. Earth Sci Rev 182:50–67
Mount J (1985) Mixed siliciclastic and carbonate sediments: a proposed first-order textural and compositional classification. Sedimentology 32:435–442
Munnecke A, Servais T, Vachard D (2000) A new family of calcareous microfossils from the silurian of Gotland, Sweden. Palaeontology 43:1153–1172
Mutterlose J, Bornemann A, Herrle J (2009) The Aptian Albian cold snap: evidence for “mid” Cretaceous icehouse interludes. N Jb Geol Paläont Abh 252:217–225
Myrow P (1992) Bypass-zone tempestite facies model and proximality trends for an ancient muddy shoreline and shelf. SEPM JSR. https://doi.org/10.1306/D426789D-2B26-11D7-8648000102C1865D
Myrow P (2005) Storms and Storm Deposits. In: Encyclopedia of Geology, Elsevier, 580–587.
Nebelsick JH, Bassi D (2000) Diversity, growth forms and taphonomy: key factors controlling the fabric of coralline algae dominated shelf carbonates. SP, 178: 89–107.
Nelson CS (1988) An introductory perspective on non-tropical shelf carbonates. Sed Geol 60:3–12
Nori L, Lathuilière B (2003) Form and environment of Gryphaea arcuata. Lethaia 36:83–96
O’Brien NR, Slatt RM (1990) Argillaceous rock atlas. Springer, New York, New York, NY
Olivero EB (1983) Amonoideos y bivalvos berriasianos de la cantera Tres Lagunas, Chubut. Ameghiniana 20:11–20
Olivero E (1987) Cefalópodos y bivalvos titonianos y hauterivianos de la Formación Lago La Plata, Chubut. Ameghiniana 24:181–202
Olivero E, Aguirre-Urreta MB (2002) Sucesión de amonoideos de la Formación Katterfeld (Valanginiano-Hauteriviano) en su área tipo, Lago Fontana, Chubut. In: XV Congr. Geol. Argentino, Actas, El Calafate, Argentina, 6 pp.
Olivero E (1982) Estratigrafía de la cuenca sur del Lago Fontana, Provincia del Chubut. PhD Thesis, Universidad de Buenos Aires
Pankhurst RJ, Weaver SD, Hervé F, Larrondo P (1999) Mesozoic-Cenozoic evolution of the North Patagonian Batholith in Aysen, southern Chile. J Geol Soc London 156:673–694
Pankhurst R, Hervé F, Fanning M, Suárez M (2003) Coeval plutonic and volcanic activity in the Patagonian Andes: the Patagonian Batholith and the Ibáñez and Divisadero formations, Aysén, Southern Chile. In: X Congr. Geol. Chileno, Actas, Concepción, Chile, 5 pp.
Pedley M, Carannante G (2006) Cool-water carbonate ramps: a review. J Geol Soc London Spec Publ 255:1–9
Pérez-López A, Pérez-Valera F (2012) Tempestite facies models for the epicontinental Triassic carbonates of the Betic Cordillera (southern Spain): tempestite models for the epicontinental Triassic, southern Spain. Sedimentology 59:646–678
Petford N, Turner P (1996) Reconnaissance 40Ar-39Ar age and palaeomagnetic study of igneous rocks around Coyhaique, S. Chile (45°30’-47°S). In: Third ISAG, St Malo, France, 625–628.
Plint AG (2010) Wave- and storm-dominated shoreline and shallow-marine systems. In: Facies models 4, 4. Version (Ed. N.P. James and R.W. Dalrymple), Geolog. Assoc. of Canada, St. Johns, 167–200.
Ploszkiewicz JV, Ramos VA (1977) Estratigrafía y tectónica de la Sierra de Payaniyeu (Provincia del Chubut). Rev. Asoc. Geol. Argentina, XXXII: 209–226.
Ploszkiewicz JV (1987) Descripción Geológica de la Hoja 47 c—“Apeleg”, Provincia del Chubut: Carta Geológico-económica de la República Argentina, Escala 1:200.000. SEGEMAR Boletín, 100 pp.
Pomar L, Brandano M, Westphal H (2004) Environmental factors influencing skeletal grain sediment associations: a critical review of Miocene examples from the western Mediterranean: tropical foramol associations. Sedimentology 51:627–651
Price G (1999) The evidence and implications of polar ice during the Mesozoic. Earth Sci Rev 48:183–210
Price GD, Ruffell AH, Jones CE, Kalin RM, Mutterlose J (2000) Isotopic evidence for temperature variation during the early Cretaceous (late Ryazanian–mid-Hauterivian). JGS 157:335–343
Price GD, Janssen NMM, Martinez M, Company M, Vandevelde JH, Grimes ST (2018) A high-resolution belemnite geochemical analysis of Early Cretaceous (Valanginian-Hauterivian) environmental and climatic perturbations. Geochem Geophys Geosyst 19:3832–3843
Price GD (2009) Mesozoic Climates. In: Encyclopedia of Earth Sciences Series: Encyclopedia of Paleoclimatology and Ancient Environments (Ed. V. Gornitz), Springer Science+Business Media, 554–558.
Pucéat E, Lécuyer C, Sheppard SMF, Dromart G, Reboulet S, Grandjean P (2003) Thermal evolution of Cretaceous Tethyan marine waters inferred from oxygen isotope composition of fish tooth enamels. Paleoceanography, 18: 7–1/7–12.
Rabenhorst L (1863) Kryptogamen-Flora von Sachsen, Ober-Lausitz, Thüringen und Nord-Böhmen, mit Berücksichtigung der benachbarten Länder: erste Abtheilung: Algen im weitesten Sinne, Leber und Laubmoose. Eduard Kummer, Leipzig, 653 pp.
Rafinesque C (1815) Analyse de la Nature ou Tableau de l’University et des Corps Organisés. Juean Barravecchia, Palermo.
Ramalho RS, Quartau R, Trenhaile AS, Mitchell NC, Woodroffe CD, Ávila SP (2013) Coastal evolution on volcanic oceanic islands: a complex interplay between volcanism, erosion, sedimentation, sea-level change and biogenic production. Earth-Sci Rev 127:140–170
Ramos VA (1978) Los arrecifes de la formación cotidiano (Jurásico Superior) en la cordillera patagónica y su significado paleoclimático. Ameghiniana 15:97–111
Ramos ME, Suárez R, Boixart G, Ghiglione M, Ramos VA (2019) The structure of the northern Austral Basin: tectonic inversion of mesozoic normal faults. J S Am Earth Sci 94:102197
Ramos VA, Lazo DG (2006) La Formación Tres Lagunas en las nacientes del arroyo Blanco al sur de lago La Plata—Chile. In: XI Congr. Geol. Chileno, Actas, Antofagasta, Chile, 2, 299–302.
Ramos VA (1976) Estratigrafía de los lagos La Plata y Fontana, Provincia del Chubut, República Argentina. In: I Congr. Geol. Chileno, Actas, Santiago, Chile, 1, 43–64.
Ramos VA (1981) Descripción Geológica de la Hoja 47 ab—“Lago Fontana”, Provincia del Chubut: Carta Geológico-económica de la República Argentina, Escala 1:200.000. SEGEMAR Boletín, 135 pp.
Riccardi A (1977) Berriasian invertebrate fauna from the Springhill Formation of southern Patagonia. N Jb Geol Paläont 155:216–252
Risk MJ, Edinger E (2011) Impacts of sediment on coral reefs. In: Hopley D (ed) Encyclopedia of Modern Coral Reefs. Springer, Netherlands, Dordrecht, pp 575–586
Rivas H, Salazar C, Stinnesbeck W (2021) Facies and sequence stratigraphy of a mixed carbonate-volcaniclastic ramp in intra-arc settings: an example from the Toqui Formation (Lower Cretaceous), southern Chile (45°S). J S Am Earth Sci 109:103292. https://doi.org/10.1016/j.jsames.2021.103292
Rubilar AE, Lazo DG (2009) Description of Aetostreon pilmatuegrossum sp. nov. from the Lower Cretaceous of Argentina (Neuquén Basin), and significance of the conservative left valve morphology in oysters of the genus Aetostreon Bayle. Cretac Res 30:727–748
Rubilar AE (2000) Aetostreon sp. nov. del Neocomiano, sur de Chile y Argentina, y su afinidad morfológica con Gryphaea Lamarck. In: IX Congr. Geol. Chileno, Actas, Puerto Varas, Chile, 2, 249–253.
Salazar C, Stinnesbeck W (2015) Redefinition, stratigraphy and facies of the Lo Valdés Formation (Upper Jurassic-Lower Cretaceous) in central Chile. Boletín Del Museo Nacional De Historia Natural 64:41–68
Scalabrino B (2009) Déformation d’un continent au-dessus d’une dorsale océanique active en subduction. PhD Thesis, Université Montpellier 2 Sciences et Techniques du Languedoc
Scasso RA, Kiessling W (2002) Earliest Cretaceous high latitude reefs in Tres Lagunas (Chubut Province, Argentina). In: XV Congr. Geol. Argentino, Actas, El Calafate, Argentina, 6 pp.
Scasso RA (1987) Estratigrafía y ambientes de sedimentación del ciclo sedimentario del Jurásico Superior y Cretácico Inferior de la región sudoccidental del Chubut, con referencias a la columna estratigráfica general del área. PhD Thesis, Universidad de Buenos Aires
Scasso RA (1989) La cuenca sedimentaria del Jurásico Superior y Cretácico Inferior de la región sudoccidental del Chubut. In: Chebli GA, Spalletti LA (eds) Cuencas Sedimentarias Argentinas, pp 395–417
Schlager W (2003) Benthic carbonate factories of the Phanerozoic. Int J Earth Sci 92:445–464
Schlager W (ed) (2005) Carbonate sedimentology and sequence stratigraphy. In: SEPM concepts in sedimentology and paleontology, vol 8. Society for Sedimentary Geology
Schneider J-L (2000) Volcaniclastic sedimentation in submarine settings: products and processes. In: Leyrit H, Bordet P, Montenat C (eds) Volcaniclastic rocks, from magmas to sediments. Gordon and Breach Science Publishers, Amsterdam, pp 175–192
Scholle PA, lmer-Scholle DS (2003) A color guide to the petrography of carbonate rocks: grains, textures, porosity, diagenesis. AAPG, Tulsa, Ok, 474 pp.
Seton M, Müller RD, Zahirovic S, Gaina C, Torsvik T, Shephard G, Talsma A, Gurnis M, Turner M, Maus S, Chandler M (2012) Global continental and ocean basin reconstructions since 200Ma. Earth-Sci Rev 113:212–270
Shanmugam G (1997) The Bouma Sequence and the turbidite mind set. Earth-Sci Rev 42:201–229
Shanmugam G (2002) Ten Turbidite Myths. Earth-Sci Rev 58:311–341
Shanmugam G (2016) Slides, slumps, debris flows, turbidity currents, hyperpycnal flows, and bottom currents. In: Encyclopedia of Ocean Sciences, Elsevier, 228–257.
Silva PC, Johansen HW (1986) A reappraisal of the order Corallinales (Rhodophyceae). Brit Phycol J 21:245–254
Simone L, Carannante G (1988) The fate of foramol (“temperate-type”) carbonate platforms. Sed Geol 60:347–354
Skarmeta J (1976) Evolución tectónica y paleogeográfica de los Andes Patagónicos de Aisén (Chile), durante el Neocomiano. In: I Congr. Geol. Chileno, Actas, Santiago, Chile, B, 1–15.
Steinmann G (1930) Sobre Archamphiroa jurassica. Revista del Museo de la Plata, 1–8.
Stenzel, H.B. (1971) Oysters. In: Treatise in Invertebrate Paleontology, Volumen 3, Mollusca 6 (Bivalvia) (Ed. R.C. Moore), Geological Society of America; University of Kansas, 3, N953–N1224.
Suárez M, De la Cruz R (1994a) Estratigrafía del Jurásico Superior - Cretácico Inferior de la Cordillera Patagónica Oriental (45°–46° latitud sur), Chile: Facies, Paleogeografía. SERNAGEOMIN, Región de Aysén, Chile, 98 pp
Suárez M, De la Cruz R (1994b) Estratigrafía y paleogeografía mesozoica de Aisén Nororiental. In: VII Congr. Geol. Chileno, Actas, Concepción, Chile, I, pp 538–542
Suárez M, De la Cruz R (2001) Jurassic to miocene K-Ar dates from eastern central Patagonian Cordillera plutons, Chile (45°–48° S). Geol Mag 138:53–66
Suárez M, De la Cruz R, Bell CM (1996) Estratigrafía de la región de Coyhaique (latitud 45°–46° S), Cordillera Patagónica, Chile. In: XIII Congr. Geol. Argentino y III Congr. Explor. Hidroc., Actas, I, pp 575–590
Suárez M, De la Cruz R, Aguirre-Urreta MB, Fanning M (2005) Diachronic Tithonian-Valanginian marine transgression of the Coyhaique Group, Aysén Basin (43°-47°S), Chile. In: XVI Congr. Geol. Argentino, Actas, La Plata, Argentina, I, 305–308.
Suárez M, De la Cruz R, Bell CM (2007) Geología del área Ñireguao-Baño Nuevo, Región Aisén del General Carlos Ibáñez del Campo. SERNAGEOMIN, Santiago, Chile, 56 pp
Suárez M, De la Cruz R, Aguirre-Urreta B, Fanning M (2009) Relationship between volcanism and marine sedimentation in northern Austral (Aisén) Basin, central Patagonia: Stratigraphic, U-Pb SHRIMP and paleontologic evidence. J S Am Earth Sci 27:309–325
Suárez M, De la Cruz R, Bell M, Demant A (2010a) Cretaceous slab segmentation in southwestern Gondwana. Geol Mag 147:193–205
Suárez M, Demant A, Cruz RDL, Fanning CM (2010b) 40Ar/39Ar and U-Pb SHRIMP dating of Aptian tuff cones in the Aisén Basin, Central Patagonian Cordillera. J S Am Earth Sci 29:731–737
Suárez M, Gressier JB, Rossel P, De la Cruz R (2023) Lower cretaceous missing volcanic arc. A migrating arc, central Patagonian cordillera, Chile: Detrital zircon U–Pb geochronology. Int J Earth Sci (Geol Rundsch) 112:907–923. https://doi.org/10.1007/s00531-022-02272-4
Swift D, Figueiredo A Jr, Freeland GL, Oertel GF (1983) Hummocky cross-stratification and megaripples: A geological double standard? SEPM JSR. https://doi.org/10.1306/212F8369-2B24-11D7-8648000102C1865D
Tarney J, Windley BF, Kröner A (1981) Marginal basins through geological time. Phil Trans r Soc Lond A 301:217–232
Tennant JP, Mannion PD, Upchurch P, Sutton MD, Price GD (2017) Biotic and environmental dynamics through the Late Jurassic-Early Cretaceous transition: evidence for protracted faunal and ecological turnover: Jurassic-Cretaceous biotic and abiotic dynamics. Biol Rev 92:776–814
Toscano AG, Lazo DG (2020) Taxonomic revision and palaeobiogeographic affinities of Berriasian-Valanginian oysters from the Vaca Muerta and Mulichinco formations, southern Mendoza, Neuquén Basin. Argentina Cretaceous Research 109:104358
Toscano AG, Lazo DG, Luci L (2018) Taphonomy and paleoecology of lower cretaceous oyster mass occurrences from west-central argentina and evolutionary paleoecology of gregariousness in oysters. Palaios 33:237–255
Townsend MJ (1998) The palaeogeography of the Lower Cretaceous Aysén Basin of southern Chile. PhD Thesis, University of Bristol
Ulmer-Scholle DS, Scholle PA, Schieber J, Raine RJ (2014) A color guide to the petrography of sandstones, siltstones, shales and associated rocks. American Association of Petroleum Geologists, Tulsa, OK, U.S.A, 526 pp.
van de Schootbrugge B, Föllmi KB, Bulot LG, Burns SJ (2000) Paleoceanographic changes during the early Cretaceous (Valanginian–Hauterivian): evidence from oxygen and carbon stable isotopes. Earth Planet Sci Lett 181:15–31
Vialov OS (1936) Sur la classification des huitres. In: Academie des Science de l’URSS, Comptes Rendus (Doklady), 13, 17–20.
Vickers ML, Bajnai D, Price GD, Linckens J, Fiebig J (2019) Southern high-latitude warmth during the Jurassic–Cretaceous: New evidence from clumped isotope thermometry. Geology 47:724–728
Walker RG, Plint AG (1992) Wave- and storm-dominated shallow marine systems. In: Facies models: response to sea level change, 8. (Ed. R.G. Walker and N.P. James), Geological Association of Canada, Toronto, 219–238.
Weissert H, Erba E (2004) Volcanism, CO2 and palaeoclimate: a Late Jurassic-Early Cretaceous carbon and oxygen isotope record. J Geol Soc 161:695–702
Westphal H, Halfar J, Freiwald A (2010) Heterozoan carbonates in subtropical to tropical settings in the present and past. Int J Earth Sci (geol Rundsch) 99:153–169
Woelkerling W, Irvine L, Harvey A (1993) Growth-forms in non-geniculate coralline red algae (Coralliinales, Rhodophyta). Aust Systematic Bot 6:277
Wray JL (1977) Calcareous algae. Elsevier Scientific Pub. Co.; Distributors for the United States and Canada, Elsevier/North Holland, Amsterdam, New York
Wright VP (1992) A revised classification of limestones. Sed Geol 76:177–185
Wright VP, Burchette TP (1996) Shallow-water carbonate environments. In: Reading HG (ed) Sedimentary environments: processes, facies, and stratigraphy, 3rd edn. Blackwell Science, Oxford, pp 325–394
Acknowledgements
We gratefully acknowledge Prof. Dr. Ioan Bucur (Babes-Bolyai University), and Dr. Stephen Kershaw (Brunel University London) for their help in the determination of microfossils. We appreciate the guidance of the Prof. Dr. Axel Munnecke (Universität Erlangen-Nürnberg), and Dr. Alexander Varychev (Universität Heidelberg) during the acquisition of SEM images. The authors would like to thank Valentina Maldonado, Benjamín Aldridge and Rayen Álvarez for their support during field work. We are grateful to Dr. Roberto Scasso (Universidad de Buenos Aires) and one anonymous reviewer for their careful revision and constructive remarks on the manuscript, and Enrique Bostelmann for his thoughtful observations.
Funding
Open Access funding enabled and organized by Projekt DEAL. Financial support to this project was provided by the Chilean National Fund for Scientific and Technological Development (FONDECYT de Iniciación 11140176), by the Chilean National Agency for Research and Development (ANID/DOCTORADO BECAS CHILE/2016–72170384), and by the German Academic Exchange Service (DAAD, STIBET/ “Studienabschlussbeihilfe Stipendium”).
Author information
Authors and Affiliations
Contributions
HR: software, formal analysis, investigation, data curation, writing—original draft, writing—review and editing, visualization. CS: conceptualization, methodology, writing—review and editing, supervision, funding acquisition. WS: resources, writing—review and editing, supervision, project administration.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no competing interests to declare that are relevant to the content of this article.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rivas, H., Salazar, C. & Stinnesbeck, W. A “cool-water”, non-tropical, mixed volcaniclastic–carbonate ramp from the Early Cretaceous of southern Chile (45°40’S). Facies 69, 14 (2023). https://doi.org/10.1007/s10347-023-00669-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10347-023-00669-4