
Abstract
We present a comprehensive data-based characterization of the subsistence economy of Chalcolithic Cucuteni–Trypillia societies (CTS) on the Moldovian and Suceava plateaus and the Podolian and the Dnieper uplands. This study is based on a quantitative evaluation of archaeobotanical samples from 34 settlement sites, with a focus on Trypillia mega-sites and on stable isotopic analysis of ancient crop residues. The isotopic analysis allows us to identify specific cultivation strategies, which show a close relationship with animal husbandry for manure. We describe the economy of the Trypillia mega-sites as having been based on an elaborate agricultural system, in which the inhabitants knew how to grow crops that could withstand the ecological constraints of growth, especially along the forest steppe ecotone. We also argue that the agglomeration of greater population densities at these mega-sites contributed to landscape change from woodland and forest to open grassland and steppe. Following on from this, we suggest that cultivation practices of the CTS were important in the establishment of the present-day cultural steppe in this region.
Similar content being viewed by others

Introduction
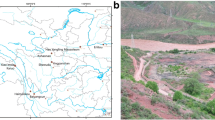
Population agglomerations of thousands of inhabitants in Chalcolithic mega-sites of the Cucuteni–Trypillia societies (CTS) occurred across the Suceava and the Moldovian plateaus and the Podolian and the Dnieper uplands. This region is in the semi-arid forest steppe ecotone (Fig. 1) and has a humid continental climate, with wet winters and warm summers (Dfb zone; Köppen and Geiger 1939). The Trypillia settlements represented a new spatial concept of social organization which spread eastwards from the Carpathian mountains around 4800 bce, with their core area in the southern river Bug region in the late 5th and early 4th millennia bce. The chronology of CTS is divided into three archaeological phases (in which Trypillia is shortened to Tryp.), early (Tryp. A-B1), middle (divided into Tryp. B1/B2 and Tryp. B2-C1) and late (Tryp. C2). The mega-sites were built in the middle phases (Tryp. B1/B2-C1), between ca. 4300/4100 and 3650 bce. The mega-site period was preceded and followed, for the most part, by dispersed small and mid-sized settlements.

Distribution of Cucuteni–Trypillia sites in the forest steppe ecotone, shown as blue dots, in sizes indicating the number of investigated features per site. Dnieper upland (1–12); 1, Grebenjukiv; 2, Chyzhivka; 3, Veselyi Kut; 4, Nebelivka; 6, Volodymyrivka; 7, Moshuriv; 8, Maidanetske*; 9, Talianky; 10, Apolianka; 11, Kosenivka; 12, Rohy. Podolian upland (5, 13–24); 5, Trostianchyk; 13, Bernashivka; 14, Hariachkivka 3; 15, Hariachkivka 7; 16, Hariachkivka 8; 17, Ternivka; 18, Vilshanka; 19, Zabolotne; 20, Bilyi Kamin; 21, Chechelnyk; 22, Viitivka; 23, Kisnytsia; 24, Krynychky-Ferma. Suceava Plateau; 25, Valeni-Cetatuie. Moldavian plateau (26–34); 26, Shcherbaky-Cuconesti Vechi; 27, Carbuna 1; 28, Brinzeni IV; 29, Ozheve; 30, Rusestii Noi; 31, Stefanesti-Hulboca; 32, Stolniceni; 33, Cunicea 1; 34, Cunicea 3, 4. Italics indicate sites from the literature, asterix (*) indicates the site with data from our own research and from the literature (from Map of the Natural Vegetation of Europe, Bohn et al. 2004; graphics by Esther Thelen, IUFG Kiel)
Previous research has shown that the decline of the Trypillia mega-sites was caused by the centralization of political power (Hofmann et al. 2019) and not by environmental factors. The soils, hydrology and vegetation should have been able to sustain this society with its subsistence economy (Dal Corso et al. 2019; Dreibrodt et al. 2020).
To add to our understanding of the rise and fall of the Trypillia mega-sites, we investigate economic, social and political changes during this period from a study of economically significant plants, through a statistical analysis of charred botanical macroremains from 34 sites, many of them Trypillia mega-sites, supplemented by stable isotope analyses of charred crop remains from some of these same sites.
The Trypillia mega-sites
Trypillia mega-sites cover 100 to 320 ha each and contained thousands of houses built in rings around a central open space, a layout that implies the intention of social congregation (Rassmann et al. 2014). The inhabitants were organised into neighbourhoods, each with a mega-structure interpreted as an assembly hall and some with pottery workshops (Chapman et al. 2016; Ohlrau 2020). Most authors suggest that the large sites were permanent settlements with populations exceeding 10,000 inhabitants (Kruts et al. 2008; Rassmann et al. 2014; Müller and Rassmann 2016; Hofmann et al. 2019; Ohlrau 2020). The social organization at these sites can be reconstructed from the remains of buildings and the finds of the house inventories. Dwellings with a standard shape and standardised inventories were spatially arranged in neighbourhoods or quarters, each of which had a slightly larger “mega-structure” that according to its specific inventory is interpreted as a communal building. This indicates a society in which decision-making processes were communally negotiated. The involvement of the whole community in decision-making seems to have ended at a certain time, when political power was centralised. In the case of the site of Maidanetske, such a communal society existed for more than 150 years and then collapsed, it is assumed due to political centralization, as the archaeological record shows the decline of medium sized mega-structures and the establishment of one large megastructure there (Müller et al. 2016, 2018; Hofmann et al. 2019). The technological innovation of animal-drawn sledges facilitated agriculture by making it possible to cover long distances in and around the mega-sites. The fields within the sites, that most probably were located a relatively long way from the dwellings and storage areas, were reached in this way, as well as those at the centre of the settlement (Shatilo 2021, pp 229–234). Animal husbandry practices involved a dual pasturing system, combining extensive pasturing of sheep, goats and some cattle outside the settlement, and also intensive keeping of cattle in the settlement for milk and meat and the rearing of pigs which were fed on household waste (Makarewicz et al. 2022).
The population agglomeration in the Trypillia mega-sites is unique for Europe at this time. They were partly contemporary with the Chalcolithic urbanization processes in northern Mesopotamia, with settlements of 10–20 ha, for example, at Tell Brak (Lawrence and Wilkinson 2015). They pre-date the second wave of urbanization in Mesopotamia in the Early Bronze Age (EBA, 2600−2000 bce), which saw settlement sizes of 40–120 ha as at Uruk (Akkermans and Schwartz 2003, p 186; Müller and Pollock 2016). Their exceptional nature raises two questions: (1) How was the subsistence economy of these Trypillia mega-sites organised to make them sustainable? And (2) What made so many people gather or permanently settle in this forest steppe region? Our data presented here suggest that the people were growing crops and raising livestock all year round and that the Trypillia mega-sites can be characterized as rural settlements, although of exceptional size and with a complex, communally organised subsistence system, in contrast to the urban settlements of Tell Brak and Uruk (Lawrence and Wilkinson 2015; Styring et al. 2022).
State of the art for Cucuteni–Trypillia (CTS) archaeobotany
CTS sites have been the subject of archaeobotanical study since 1930 (see discussions in Pashkevych and Videiko 2006; Kirleis and Dal Corso 2016). In earlier analyses, due the focus on house structures by the field archaeologists, early data on the plant economy came from plants used as temper in daub, accidental impressions of plant remains on potsherds and mass finds of plant remains in vessel fills. These studies identified the main crops as the cereals Triticum monococcum (einkorn), T. dicoccum (emmer) and Hordeum vulgare (barley), as well as pulses such as Pisum sativum (pea) and Vicia ervilia (bitter vetch). Single finds of Triticum aestivum (bread wheat), cf. T. spelta (possibly spelt) and Lens culinaris (lentil) broadened the crop spectrum. The drawback of plant impressions is that they are hard to identify and that their representativeness is difficult to judge (An et al. 2019; Endo et al. 2022). Their advantage is that they reveal the presence of plants that are rarely found in assemblages of charred material, such as ones which were eaten raw and soft tissues from leaves (Videiko 2017). Thus, the gathering of wild fruits from the native woodlands is shown by imprints of whole fruits, seeds, pips and fruitstones of apple Malus (apple), pear Pyrus (pear), Prunus avium (cherry), P. spinosa (sloe), Cornus mas (cornelian cherry), Crataegus (hawthorn), Viburnum lantana (wayfaring tree), Sambucus ebulus (dwarf elder), Solanum nigrum (black nightshade) and Corylus avellana (hazel), as well as acorns of Quercus (oak). Imprints of rather large grape pips indicate the use of Vitis vinifera ssp. sylvestris, (wild grape) which is native to Ukraine. Further imprints, some of which may need critical reassessment, were identified as Prunus armeniaca (apricot), P. insititia (damson) and P. cerasifera (cherry plum) (Yanushevich 1975, 1978, 1980; Körber-Grohne 1996, pp 181, 211; Pashkevych and Videiko 2006, p 41; Zohary et al. 2012; Gluza 2013).
Archaeobotanical investigation of soil samples from the Trypillia sites of Bernashivka, Ozheve, Nebelivka and Talianki has recently begun (Kruts et al. 2008, 2009; Pashkevych 2014; Chapman et al. 2020), as well as in our own work on Maidanetske, where in the 2013 excavations (Müller et al. 2017), systematic sampling for archaeobotanical remains covered various archaeological contexts, not only house structures, the larger of which included mega-structures, but also pits, kilns, fireplaces, ditches, occupational layers, levelling layers, topsoil and buried soil. The systematic sampling of different archaeological contexts in the settlements has enabled us to secure charred archaeobotanical material, which represents a sound basis for evaluating the role of plants in the CTS economy, and is complemented by the results from find concentrations such as from vessel fills, impressions in potsherds and temper in daub. Our archaeobotanical investigations were hampered by the fact that archaeologists traditionally concentrated on house structures, with masses of often burnt daub from house construction, and distribution of artefacts and clay structures in situ (for example, Chernovol 2012; Müller et al. 2017; Rud et al. 2019). Since the remains of house structures are preserved close to the modern land surface, the preservation conditions for archaeobotanical assemblages relating to these structures are far from ideal and they yield only very few, often poorly preserved plant remains, and in some cases analysis is further complicated by inclusion of modern charred material from the burning of straw and chaff on the fields after the harvest. Bucket flotation using sieves of 0.3 mm mesh led to the recovery, for the first time at any of the CTS sites, of tiny plant remains, such as cereal by-products, weed seeds, fruits and tiny awn fragments of Stipa (feather grass). This refinement of recovery methods has allowed us to distinguish various different plant-related activities and their spatial organization on the site. Stable isotopic analyses of the charred plant remains have revealed specific cultivation practices and environmental conditions, and thus can hint at climatic variability.
Material, methods and radiocarbon dating
Sampling strategy, taphonomy and sample preparation
Archaeobotanical analyses on sites of the CTS have increased considerably in the past 10 years in the number of investigated sites (Fig. 1; ESM 1, 2). Our desire to research economic questions led us to change our excavation strategy to one that concentrated on pits associated with houses, since most pits reflect the spatial patterns of the houses within the site (Rassmann et al. 2014, p 132) and a combination of house and pit (or house pit) is considered to be representative of a household as a socio-economic unit on a mega-site (Müller et al. 2016). Therefore, the finds from these pits, which contain secondary fills, should reflect the activities of these households, including remains from food processing, preparation and storage. This sampling by house pits improves archaeobotanical recovery in particular, as these pits, which go deep down into the loess soil substrate, offer very good preservation conditions for ancient charred plant remains and are usually undisturbed by modern farming. The pits turned out to be the best archive for investigation of the consumption, production, specialization and hierarchy of the socio-economic unit of a household (Dal Corso et al. 2018). The relatively high find concentrations compared with other archaeological contexts have already been documented, as for the sites Maidanetske (Dal Corso and Kirleis in press), Bilyi Kamin and Kisnytsia in Ukraine and the site Stolniceni in Moldova. The potential of these pits for Trypillia research has been described in detail by Terna (2021, pp 42–45, 55), who invented “pit archaeology” and interprets “houses as snapshots and pits as durable narratives”.
We undertook careful bucket flotation with fine mesh sieves of 0.3 mm in order to optimize our chances of recovering even tiny and fragile plant remains (Dal Corso et al. 2019; Dal Corso and Kirleis in press). We sorted all the plant remains with an Olympus SZ51 stereomicroscope at 8–40× magnification and identified them with help of the literature, such as Bojňanský and Fargošová (2007), and Cappers et al. (2012), and the large reference collection of seeds and fruits at the Institute for Prehistoric and Protohistoric Archaeology in Kiel.
The dataset
Our dataset includes charred plant remains from the 23 sites for which we undertaken flotation and analysis, supplemented by published data for another 11 sites, in all 34 sites (with Maidanetske represented in both categories) (ESM 1). We restricted comparisons between sites to the 16 sites with more than ten cereal grains each. The remaining 18 sites have small plant assemblages, with between zero and five cereal grains; some of these are mass finds, often vessel fills, of non-cereals such as Pisum. For most sites, we considered 1–10 features, yielding a total of 1–30 samples. For two sites, we considered many more features and samples, as from Stolniceni (map site identifier 32) with 289 samples from 92 features and Maidanetske (8) with 752 samples from 162 features (Fig. 1).
Of the 34 sites, seven date to the early phase Cucuteni (Cuc.) A, Tryp. A, B1, 4750−4300/4100 bce) slightly preceding the mega-sites phenomenon of Trypillia (Tryp.) B1/B2, 4300/4100−3900/3800 bce); 13 belong to the middle phase, the mega-site phenomenon (Tryp. B2, C1, 3900/3800−3650 bce) and another seven belong to the late phase (Tryp. C2, 3650−3000 bce), with dispersed settlements. The site chronology is intertwined with the geographical site distribution, which follows a west-east gradient, with the early sites in the west and the large mega-sites (> 100 ha) in the Dnieper upland, in the east.
Overall, we recorded 21,550 charred plant remains from the 34 sites, including indeterminates and estimates for abundance and ubiquity in data from publications, with 10,361 seeds and fruits in all. Note that for the sites marked with * in ESM 1, the number of finds had to be estimated based on the semi-quantitative information available in the literature. For three of the sites where flotation was used, the Kiel team identified an additional 3,505 mineralized seeds and fruits, dominated by a mass find concentration of Chenopods from Maidanetske.
Correspondence analysis (CA) was carried out with CAPCA v. 3.1 add-in for Microsoft Excel and boxplots were created with PAST v. 4.11 (Hammer et al. 2001; Madsen 2016). For CA, we considered all the cereal grains and by-products from archaeological sites where more than ten cereal grains were recovered.
14C dates on charred plant remains
We directly AMS dated remains of the main crops and some other plant taxa retrieved from Cucuteni–Trypillia sites (Fig. 2, ESM 3). Direct radiocarbon dating of plant remains is important to confirm (or, indeed, disprove) whether they are the same date as the archaeological context of recovery, especially if another chronology might be expected (for example, Grikpėdis and Motuzaite Matuzeviciute 2018; Golea et al. 2023), especially for remains prone to being intrusive such as Panicum (millet) grains (Filipović et al. 2020), as shown by ones from Maidanetske that gave a date in the 1st millenium ce (Dal Corso et al. 2022; ESM 3). Conversely, 14 C dating of tiny awn fragments of the steppe plant Stipa (feather grass) from Maidanetske and Stolniceni ruled out that they had moved down from later layers through bioturbation (Fig. 2). Single grains of hulled barley from Maidanetske were revealed to be recent, but Hordeum vulgare var nudum (naked barley) was shown to be contemporary with the site (Fig. 2; ESM 3). The hulled wheats Triticum dicoccum (emmer), T. monococcum (einkorn) and T. timopheevii (Timopheev’s wheat) and also Pisum sativum (pea), Corylus (hazelnut), Chenopodium album (goosefoot) and Sambucus ebulus (dwarf elder) were identified from Trypillia. The 14C dating programme allowed us to confirm that most of the plant remains are contemporary with the sites where they were retrieved and thus contribute to the clarification of the site chronologies.

Radiocarbon dates calibrated using the Intcal20 (Reimer et al. 2020) calibration curve and Oxcal v. 4.4, function R_Date (Bronk Ramsey 2009). Bars indicate 95.4% probability intervals, plus symbols indicate median ages. The samples come from the Tryp. B1 site of Chyzhivka (CHI); the Tryp. B2/C1 sites Stolniceni (STO), Chechelnyk (CHE) and Bilyi Kamin (BiK); the Tryp. C1 site Maidanetske (MAID); and the Tryp. C2 sites Kosenivka (KOS) and Cunicea (CUN). Sample taxonomic abbreviations: n-barley, naked barley; ngw, new glume wheat syn. Triticum timopheevii; Chenopod, goosefoot; d-elder, dwarf elder (graphic by Carsten Reckweg, IUFG Kiel)
Stable C and N isotopes of cereals and pulses
Paired measurements of the carbon and nitrogen isotopes δ13C and δ15N on 96 samples of botanical remains have been done for the Trypillia sites Maidanetske, Stolniceni, Chechelnyk and Bilyi Kamin, and the later Trypillia C2 site of Kosenivka (ESM 4). Most of the measurements were on single cereal grains (n = 77) of emmer, einkorn, naked barley or T. timopheevii, and a further 19 on peas. Photographs of all seeds and fruits are stored in the LandMan portal of CRC1266, a database of research at Kiel university. For stable carbon isotope analyses on plants, the fractionation during photosynthesis has to be considered to allow for comparison between data from different sites. We calculated the degree of 13C fractionation, as expressed in Δ13C values, based on an atmospheric δ13C value of 6.38‰ for the time of Trypillia (Ferrio et al. 2005). Due to the low and inconsistently reported influence of charring, no δ13C corrections were made. δ15N results were moderately corrected (− 0.5‰) for comparisons with uncharred material (Poole et al. 2002; Heaton et al. 2009; Fraser et al. 2013; Nitsch et al. 2015; Philippsen et al. 2019). Unless otherwise stated, δ15N values in the text and figures represent adjusted values.
Fourier-transform infrared (FTIR) spectroscopy pre-treatment on selected finds (two peas and 13 cereal grains) was carried out at the Archaeological Stable Isotope Laboratory in Kiel. The pulses revealed no probable trace of contamination. The cereal grains showed the presence of carbonates and were therefore processed for stable isotope measurements following Vaiglova et al. (2014), at the Department of Earth and Environmental Sciences of the Katholieke Universiteit Leuven, Belgium. The charred plant material was analysed for stable isotopes through combustion using an elemental analyser (Thermo Flash HT/EA or EA 1110) coupled to an IRMS system (Thermo Delta V Advantage) via a Conflo IV interface from Thermo with the isotope standards IAEA600 (δ13C − 27.77, δ15N 1‰), Tuna (δ13C − 18.72‰, δ15N 13.77‰) and Leucine (δ13C − 13.73‰, δ15N 1.07‰). The standard deviations on measured isotope values are typically below 0.1‰. Isotope δ values are expressed relative to the international Vienna PeeDee Belemnite (VPDB) standard for 13C and AIR for 15N.
Results and discussion
Trypillia mega-sites as drivers for the expansion of grasslands
The forest steppe ecotone, a transitional zone that is particularly sensitive to alteration, has archaeological traces of human impact going back to the Palaeolithic and continuing into the Mesolithic and Neolithic, which intensified considerably in the Chalcolithic, with the establishment of the Trypillia mega-sites (Dreibrodt et al. 2020, Table 2; Saile 2020). The charcoal record, as well as wood imprints in daub used in house construction, prove the existence of patches of deciduous woodland in the vicinity of the CTS sites, showing the use of ash and oak in particular, from the riverside woodlands and from forest stands on the slopes and plateaus (Dal Corso et al. 2019). Studies of the soils at Maidanetske have revealed that the site was established on cambisol formed under woodland which was converted into chernozem (black soil) during occupation of the site. The cultivation methods of the Trypillia farmers stimulated the formation of this dark steppe soil by encouraging the digging and casting activity of anecic earthworms at Maidanetske and began the formation of the modern cultivated steppe landscape (Dreibrodt et al. 2022).
This steppe formation is confirmed by the macrobotanical record of charred Stipa (feather grass) awns from the CTS sites, the percentages of which can be used as proxy evidence for steppe expansion, and Stipa percentages increase steadily from the earliest to the latest Trypillia stage, C2 (Fig. 3). Although the recovery of Stipa remains is better when using bucket flotation with fine 0.3 mm sieves, inadequate recovery methods are not the only reason why it may be absent from the assemblage from a particular site. The areas where it occurs seem to correlate with closeness to the southern steppe region (Fig. 4). The fact that the awns are charred may indicate management of open grasslands with fire in order to expand land available for arable fields or pasture, as is known from ethnographic sources (Boivin and Crowther 2021).

Boxplot diagram showing charred Stipa (feather grass) awn fragments from the 17 sites as a proportion of all charred finds per site. Site numbers as in Fig. 1. (graphic by Carsten Reckweg, IUFG Kiel)

Charred macroremains from Trypillia organized into plant groups for the 16 sites with > 10 cereal grains. Site dates are shown by triangle, Early Tryp. (A-B1) (4750−4300/4100 bce); starburst circles, Middle Tryp. preceding mega-sites (B1/B2, 4300/4100−3900/3800 bce); solid dots, Middle Tryp. with mega-sites (B2, C1, 3900/3800−3650 bce); diamonds, Late Tryp. (C2, 3650−3000 bce). Site names and numbers as in Fig. 1 and ESM 1 (graphic by Esther Thelen, IUFG Kiel)
General overview of the Trypillia plant economy
The plant economy of the Cucuteni–Trypillia sites was, in general, mainly based on cereals, pulses and gathered plants (Fig. 4; ESM 1), see overviews in Pashkevych and Videiko (2006) and Kirleis and Dal Corso (2016).
The main plant components in the diet of the Trypillia people were cereals (Fig. 5a, b). Emmer dominates in our results and is followed in numerical importance by barley. For some barley grains, the analyst was able to distinguish between Hordeum vulgare var nudum free-threshing barley and H. vulgare var vulgare (hulled barley), thus providing confirmation that both varieties were present in the assemblages. However, 14C dating indicates that the hulled barley grains, which were present as isolated finds, are modern intrusions. Triticum monococcum (einkorn) is the second most important hulled wheat species and T. timopheevii (Fig. 6) was also part of the Trypillia cereal spectrum, possibly grown as a mixed crop together with einkorn and/or emmer. The first identification of T. timopheevii for a Trypillia context came from the Stolniceni site, and single glume bases have since been identified from the Tryp. B1/B2 sites Hariachkivka 7 and 8 (Filipović et al. in review).

Pie diagram showing percentage representation at Trypillia sites of a, cereal grains (n = 2,935 for all 34 sites); b, cereal by-products (n = 5,016 for all 34 sites). (Graphic by Carsten Reckweg, IUFG Kiel)

Charred spikelet forks of Triticum timopheevii from the mega-site (Feature 21,009, find ID 211,366) from Stolniceni, Moldova; scale bars, 1 mm (photos by Agnes Heitmann and Wiebke Kirleis, IUFG Kiel)
Remains of chaff from cereal processing have also been identified, almost exclusively of hulled wheat, with the glume bases mainly deposited in the pits accompanying the houses. The cereal by-products which could be identified to genus or species were mainly einkorn followed by emmer. The cereal grains showed the opposite ratio. However, because the proportion of unidentifiable threshing remains is very high at 51%, we need to consider the possibility that the overall proportions of the threshing remains may be closer to those of the grains themselves. Barley rachis fragments are almost completely absent, presumably because hulled wheat and free-threshing barley were treated differently after harvesting. The early processing steps for barley, which would have resulted in the removal of the rachis segments, probably occurred outside the settlement, possibly near the cornfields, as is common practice for all the free-threshing cereals (Hillman 1981; van der Veen and Jones 2006). Therefore, the barley would have been free of rachis fragments by the time it was stored in the settlement. In contrast, the early processing steps for hulled wheats resulted in the grains remaining protected by the glumes. Thus, the hulled wheats were most probably stored still articulated in their spikelets. Based on the waste from the pits, we think that de-husking of hulled wheats took place within the settlement, possibly on a day-to-day basis in each household and probably to some extent as a collective activity in the mega-structures as at Stolniceni and Kisnytsia.
For the CTS, the charred mass finds of pulses from vessel fills from Carbuna 1 (Vicia ervilia, bitter vetch, Tryp. A) and Valeni-Cetatuie (Pisum, pea Cuc. B1/Tryp. C1), both Romania, and Maidanetske (Pisum, Tryp. C1), indicate that pulses were part of the diet (ESM 1). However, most archaeobotanists believe that pulses are, in general, under-represented in samples obtained through systematic sampling (Fig. 4), because some of them can be consumed green, while others are processed for food preparation by removal of the seed coat and leaching without using fire and they may also be used to feed livestock (Filatova 2020). Thus, they do not necessarily contribute to charred plant assemblages from the economic unit of a household. From the Neolithic in south-eastern Europe, for example, the type of storage facility used is known to influence the survival of remains of certain plants in the archaeobotanical record (Filipović et al. 2018). The fact that the CTS pulses were stored in small pots fits with ethnographic observations, for example from Iberia. It may reflect cultivation of pulses in small, garden-like areas resulting in low yields. It is also possible that pulses were stored in containers made of perishable materials (Tarongi et al. 2020). Our stable isotope results such as those for Maidanetske, for example, show that small-scale legume cultivation is likely to have involved intensive cultivation strategies, with the addition of animal manure, as described below.
Perspective on the Trypillia plant economy through time
Overall, we see the usual Late Neolithic crop spectrum at the CTS sites, consisting of hulled wheat and barley, with a noteworthy component of Triticum timopheevii at the Stolniceni site. Despite the development of society shown by the transition from dispersed to agglomerated populations and back again, the spectrum of cereals cultivated by CTS people remains, overall, quite conservative.
When we look at individual sites, we see that the record is more heterogeneous in terms of the relative amounts of different cereals, although we cannot exclude the possibility that this picture has been influenced by formation processes of the archaeological record, sampling strategies and extraction methods applied (taphonomy). The records of the individual sites (Fig. 7a, b) show that emmer is a strong crop component at the mega-sites of Maidanetske (site 8), Nebelivka (4) and Kosenivka (11), the latter being a site of 80 ha at the transition to Tryp. C2. The importance of emmer in times of population agglomeration may relate to its yield, which is much higher per plant and unit area than that of einkorn, making it possible to feed many people (Miedaner and Longin 2017, pp 26–27; Kreuz 2012, p 56). Looking into the records through time, we see that after the collapse of the mega-sites, when a dispersed settlement system was established einkorn increased, while barley decreased (Figs. 8 and 9). The decrease is particularly evident at the Tryp. C2 site of Kisnytsia (23; Fig. 7a, b) in both grains and by-products, although einkorn was important as well at the earlier Tryp. B1/B2 site of Ternivka (17; Fig. 7a) and the early Trypillia site of Bernashivka (13; Fig. 7a). It remains to be seen whether this development is an indication of changes in culinary practices, a change in storage techniques or re-organization of society into dispersed settlements. The fact that einkorn was valued in the later Trypillia dispersed settlements (Tryp. C2) may be due to its tough stalks being increasingly used for binding or thatching. Moreover, being a robust plant, it could have ensured the yield in mixed cropping. Growing einkorn with emmer helps the emmer stalks stay upright under conditions of heavy rain (Kreuz 2012, p 56).

Pie diagrams showing numbers of remains from the 16 sites with > 10 cereal grains; A, cereal grains; B, cereal by-products. Open circles, sites with > 10 cereal grains with no by-products recorded; triangles, Early Tryp. (A-B1) (4750−4300/4100 bce; starburst circle, Middle Tryp. preceding mega-sites (B1/B2, 4300/4100−3900/3800 bce); solid circles, Middle Tryp. with mega-sites (B2, C1, 3900/3800−3650 bce); diamonds, Late Tryp. (C2, 3650−3000 bce). Site names and numbers as in Fig. 1 and ESM 1 (graphic by Esther Thelen, IUFG Kiel)

Bar diagram showing absolute numbers and proportions of cereal grains per phase (n = 2,935 for all 34 sites combined) (graphic by Carsten Reckweg, IUFG Kiel)

Absolute numbers and proportions of cereal by-products per phase (n = 5016 for all 34 sites combined) (graphic by Carsten Reckweg, IUFG Kiel)
Spatial perspective on the Trypillia plant economy
Mega-sites exceeding 100 ha are restricted to the Dnieper upland, where the plant economy shows a correlation with the occurrence of hulled wheats, but not with barley. This spatial pattern in crop preferences is deeply intertwined with the chronological change with decreasing amounts of barley over time (Fig. 10). At the level of the individual settlements, however, a more diverse and complex picture emerges (Fig. 7a, b), but it is difficult to know whether this site-specific diversity in crop spectra is due to cultural preferences, the growing requirements of the crops, or sampling bias. The diversity seen in the genetic makeup of individual CTS human remains (Mathieson et al. 2018; Immel et al. 2020) indicates that the populations at these sites included people with a number of different geographical origins, which may have been expressed as specific culinary traditions. In a geographical sense, we see a tendency for the earlier western sites of Bernashivka (site 13), Shcherbaky (26) and Ozheve (29) to show diverse cereal assemblages, with barley and the hulled wheats emmer and einkorn. We see a similar tendency at the early ring-shaped settlements of Hariachkivka 7, 8 (15, 16) and Ternivka (17), in the Podolian upland (4300−4000 bce, Rud et al. 2022), and in the Tryp. B2 and B2/C1 sites spanning the period 4000−3800 bce (Nebelivka (4), Trostianchyk (5), Bilyi Kamin (20), Stolniceni (32) and Brinzeni IV (28). All of these sites show quite diverse crop assemblages, with emmer, einkorn, Triticum timopheevii and barley. At Maidanetske (8), with its more than 150 years of occupation, the population peak in phase 3 is expressed through a high abundance of emmer finds, which clearly dominate the record.

Correspondence analysis of cereal grain and by-products data. from sites with > 10 grains; axis 1, eigenvalues 8, explanation 32%; axis 2 eigenvalues 6, explanation 26%; combination of variables (grains and by-products) and objects (archaeological sites on an east-west transect); option “Automatic weighting of objects”. GB, glume base; RS, rachis segment (graphic by Carsten Reckweg, IUFG Kiel)
The data for the west-central Trypillia A and B1 sites Bernashivka (13), Shcherbaky (26) and Ozheve (29), as well as for the later sites Brinzeni IV (28), Nebelivka (4), Chechelnyk (21) and Kosenivka (11), come from publications of investigations that often predate systematic archaeobotanical sampling and fine-mesh sieving. That is why all three datasets are dominated by a relatively large abundance of cereal grains and include hardly any other plant groups (Fig. 4). In contrast, in sites where “pit archaeology” was applied, cereal processing by-products occur in massive quantities, together with tiny amounts of remains from gathered plants (here mainly chenopods), independently of the number of samples analysed. The rich plant assemblages from the Trypillia sites (Fig. 4) thus reflect recent methodological progress. There are two exceptions that are not as diverse, the assemblages from the mega-site of Maidanetske (site 8; Dal Corso and Kirleis in press), with its low proportions of cereal by-products and massive indication for chenopods as a gathered plant, and the site of Bilyi Kamin (20; Rud et al. 2019), with its record dominated by cereal grains. For Maidanetske, however, where many archaeological contexts were sampled, the picture changes when only the pits are considered. From the pits, cereal grains and remains of threshing residue are almost equal in number, with a slightly higher number of grains. The presence of cereal threshing remains at Maidanetske (8) is further proven through phytolith analyses on deposits from pits and temper in daub (Dal Corso et al. 2019, Fig. 6). The over-representation of cereal grains at Bilyi Kamin can most easily be explained by the fact that barley dominates the cereal spectrum (Fig. 7a) and it is commonly stored as grain only. Environmental constraints on the choice of different crops seem a less likely explanation if we consider that among the crops from neighbouring sites such as Bilyi Kamin (20), barley dominates, while at Chechelnyk (21), emmer dominates. The latter, however, is a mass find from a vessel fill (Pashkevych and Kosakivskyi 1991) and is therefore to be regarded as exceptional.
Stable nitrogen and carbon isotopes in cereals and pulses
In a forest steppe ecotone, where there are no natural deposits to provide a well dated climate reconstruction, it is difficult to draw conclusions about past cultivation conditions. Plant remains from archaeological excavations can help here, in that measurements of their stable carbon isotopes provide information about the hydrology of the soil and can thus provide clues about the past climate. For the CTS sites overall, and for each individual site, the Δ13C values and means of emmer grains (min. 15.3-mean 17.1-max. 18.5‰) and einkorn (15.9–17.4–19.9‰) are in the same range. The higher 13C depletion seen in barley (17.6–18.7–20.0‰; Fig. 11; ESM 4) can most easily be explained by its higher efficiency of water use (Ferrio et al. 2005). The 1.5‰ higher 13 C depletion in naked barley matches that from cultivation experiments with it (Kanstrup et al. 2011). The Δ13C values from CTS plant remains indicate damp climatic conditions during filling of the cereal ears, which probably occurred in late spring to midsummer, a timing that matches that of modern-day naked barley, and they fall within the range of well-watered cereals (Flohr et al. 2011). The Δ13C of the peas (17.1–18.5–19.6‰) also seems to reflect good water availability, as it is comparable to that of pulses from well-irrigated fields (Wallace et al. 2013). However, such comparisons with results from the arid environments of southwest Asia should be taken with a degree of caution, but may help to overcome the lack of a local Δ13C baseline. The Δ13C range of the peas is almost identical to that of barley, an observation already reported by Mora-González et al. (2019), and is indicative of a comparably good water use efficiency of Pisum sativum or a good water supply, possibly relating to the high water storage capacity of the humus-rich soils. So far there is no sign that the fields were irrigated. A canopy effect can most likely be ruled out as a reason for the Δ13C values of the CTS plant remains, as they were cultivated in close connection to the settlements and situated in a very open forest steppe. Even within the settlements there was sufficient open land for growing crops, as shown by the occurrence of certain snail species, as at Maidanetske (Schlütz et al. in press).

Stable isotope values of cereals and peas from Trypillia mega-sites, going from west to east. A Δ13C; B δ15N corrected for charring. The sites are Stolniceni (32), Chechelnyk (21), Bilyi Kamin (20), Maidanetske (8) and Kosenivka (11). Diamonds, Triticum dicoccum; squares, T. monococcum; crosses, T. timopheevii (NGW); triangles, H. nudum; circles, Pisum sativum. Details in ESM 4 and the CRC1266 LandMan portal at Kiel university
We can use the stable carbon values not only to understand the growing conditions of individual plants, but also to reconstruct the climate as it relates to CTS sites. The Δ13C values for all taxa suggest that farming was not affected by harsh droughts, and those from cereal grains and peas indicate a natural hydrological regime that was fully suitable for crop cultivation.
We can use the stable nitrogen isotope values to understand the cultivation methods. The δ15N values of plants show the availability of nitrogen in the soil and atmosphere for the production of plant biomass. The δ15N values of the CTS cereal grains cover a quite wide spectrum. For the hulled wheats, values corrected for charring range between 3.1 and 11.3‰ (emmer 3.4–7.6–11.3‰; einkorn 3.1–7.9–10.4‰; and T. timopheevii 3.24‰), while for naked barley, the range is narrower (6.1–7.3–7.9‰).
All of the mean values are above the 6‰ threshold argued by Bogaard et al. (2013) to indicate manuring and thus would indicate high manuring intensity. The minimum values are below this threshold and would indicate low manuring intensity. However, due to the isotope results from local palaeosoils described in Dreibrodt et al. (2022), we argue for the need to investigate whether the fertile soils in the CTS area have naturally higher stable nitrogen baseline values, in which case higher threshold values for manuring would need to be established. Also, the two soils that predate agriculture (Dreibrodt et al. 2022) show δ15N values of around 7‰, as known from natural loess soils in the Carpathian basin (Pötter et al. 2021). Nevertheless, the difference between δ15N in the soil and in plants is influenced by several factors and is difficult to estimate without local cultivation experiments. However, establishing a local geological map of isotope distribution (isoscape) and seasonal baseline is beyond the scope of this paper. The δ15N values of the three peas (Pisum sativum) from Stolniceni are low (0.7–1.1–1.7‰), while those of the 16 examples from the Maidanetske vessel fill (8) are significantly higher (2.8–5.6–7.0‰). Since pulses, including pea, meet their nitrogen requirements by fixing it from the air with symbiotic bacteria (atmosphere δ15N=0‰), manuring barely increases the δ15N in their tissues. Under the same manuring practice, pulse δ15N values can be more than 5‰ lower than those of cereal grains (Fraser et al. 2011). Therefore, even low pulse values of about 1‰ δ15N can indicate manuring (Nitsch et al. 2015). The highest values reported for modern pulses are 3–5‰ δ15N for Vicia faba (broad bean) grown on “dung soils” in Greece (Fraser et al. 2011). Such soils are a labour-intensive, specific type of anthrosol (soil resulting from human activity) that develops under long-term manuring and is often associated with dwellings and/or stables. Beans grown on a mixture of sheep dung and decomposed leaf litter during a short-term cultivation experiment showed δ15N values of 3‰ maximum for individual beans (Treasure et al. 2016). As the example of Maidanetske shows, the fertiliser available in the large settlements probably consisted of dung from cattle and pigs that may have been kept in the central square of the settlement (Makarewicz et al. 2022). Human faeces and household waste were also available as possible fertiliser for garden-like cultivated areas within the settlement. Against this background, it seems quite plausible that the peas and possibly some of the cereals retrieved from Maidanetske were grown in such small garden-like areas within the settlement or nearby, with intensive use of labour and manuring resembling that described for the formation of terra preta (Amazonian dark earth) and anthropogenic dark earth (Glaser et al. 2001; Wiedner et al. 2015). However, archaeologists have not yet identified garden or field structures at Trypillia sites.
Overall, the CTS δ15N values provide proof of crop cultivation on nutrient-rich soils within the settlement, probably created with the aid of targeted application of animal dung. Many different arable practices were used, combining extensive cereal cultivation outside the settlement on arable plots requiring relatively little labour investment, either close to or further away from the settlement, but within walking or sledge distance, and within the settlement, intensive gardening of legumes with generous manuring. Depending on the number of people to be fed, there might have been differences between the sites. For instance, for the mega-site of Maidanetske (200 ha) the δ15N of emmer grains is above 6‰ and of peas mostly above 4‰, while for the smaller site of Stolniceni (80 ha), half of the emmer δ15N was below 6‰ and peas even below 2‰.
Urban or rural? Characterising Trypillia mega-sites
The Chalcolithic Trypillia mega-sites seem to have been somewhere between farming and urban communities (Ohlrau 2022), with the extraordinary urban scale of their population agglomeration affecting both their subsistence economy and environment. A comparison of these sites with Neolithic to Bronze or Iron Age urban sites in Mesopotamia, the Aegean and southwestern Germany indicates that both the density and the scale of occupation shaped the agricultural and ecological course of development of these early cities. In terms of agricultural strategies, high density urban settlement in northern Mesopotamia and the Aegean entailed an extensification going along with a radical increase in land use, while low-density urbanism in south-western Germany allowed for more intensive management of arable land (Styring et al. 2022). However, both strategies, sometimes intertwined, are known to have been practiced in urban contexts continuously up to early modern times. Often, the hinterland served to provide economic support for feeding the large population, and farming in this hinterland was supplemented by small-scale growing within the urban space (for example, Fischer 2021, pp 115–126). Ever since populations have gathered in large agglomerations, urban sites have developed various solutions for producing plant foods. For the Trypillia mega-sites, we have been able to reveal that a broad spectrum of agricultural practices was the foundation for a complex and sustainable subsistence economy, which ranged from intensive labour investment in crop growing and animal husbandry partly within the site, and extensive crop growing and animal herding outside it. On this basis, we can define the Trypillia mega-sites as early urban sites that were deeply rooted in complex rural arrangements.
Conclusions
After decades in which knowledge of plant use at Cucuteni–Trypillia sites was mainly based on plant impressions on pottery and opportunistic finds of vessel fills, the state of research on the CTS plant economy has been significantly improved by ten years of intensive archaeobotanical investigations using a systematic sampling strategy, the shift to “pit archaeology”, and with bucket flotation using fine sieves. These measures have expanded archaeobotanical recovery to include cereal processing by-products, weeds and steppe components, in the form of Stipa (feather grass) awns. We choose to remain critical when making statements here about the economy of CTS sites, because of the heterogeneity of the archaeobotanical datasets. The data come from two sites that were intensively and systematically studied, others where up to ten 10 features were systematically studied, and still others with only incidental finds of vessel fills. Bearing this caution in mind, we are now in a position to provide a solid overview of the plant component of the CTS economy, and by including data on stable isotopes in plants, we are able to characterize the economic basis and palaeoecology of these mega-sites.
The CTS archaeobotanical record allows us to reconstruct the diachronic development of cultivation in the forest steppe ecotone of this part of eastern Europe. Agricultural expansion started with the growing of barley, emmer and einkorn together with some Triticum timopheevii at the pre-mega-site stage of the CTS (4750−4300/4100 bce; then there were changes towards consolidation of the diverse cereal spectrum, including intensive emmer and pulse cultivation, during the time of the mega-sites (3900/3800−3650 bce), and finally, more einkorn and less barley in the late Trypillia dispersed sites (3650−3000 bce). The most striking transformation in the crop spectrum is observed after the collapse of the mega-sites, when a social re-organization into dispersed settlements took place, although altogether the composition of the cereal assemblage remains conservative.
The farming and land use of the population agglomerations in the Trypillia sites in the forest steppe ecotone left its effects there and started the establishment of the present-day cultural steppe of this region. Cultivation practices promoted increased earthworm activity and encouraged the spread of Stipa and its burning, as shown by numerous charred awns dispersed in the deposits, may have been part of a grassland management system to maintain an open landscape in CTS times. From an archaeobotanical point of view, the Trypillia mega-sites can be characterized as a phenomenon that brought together diverse traditions of farming, society and culture with opportunities for knowledge transfer between what had previously been smaller communities that would have benefited from this convergence. CTS crops were cultivated on a large scale, on nutrient rich soils within a distance that could be reached by sledge from the settlement, and intensively, in garden-like areas within the settlement, in particular the latter with targeted use of animal dung. Intensive practices of growing crops were supplemented by less labour intensive growing of emmer, the latter particularly at the western mega-sites. The organization of labour at Trypillia mega-sites for access to land within and outside the settlements seems to have been done communally, negotiated between the individual households. In addition, we can assume that communal activities involved several households for integrative practices such as cereal processing. Crop growing by households would have needed agreement between them, perhaps negotiated within the built environment of the mega-structures that are interpreted as possible assembly halls. Collective cereal processing thus promoted social coherence. Overall, crop production at the Trypillia mega-sites was sustained by a complex subsistence regime in which crop cultivation and animal husbandry were deeply intertwined.
References
Akkermans PMMG, Schwartz GM (2003) The archaeology of Syria: from Complex Hunter-Gatherers to Early Urban Societies (ca. 16,000–300 BC). Cambridge University Press, Cambridge
An T, Pashkevich G, Jones M (2019) Re-examining millet impressions in Usatovo clay materials from NW Black Sea region, Ukraine. Archaeol Anthropol Sci 11:3201–3211. https://doi.org/10.1007/s12520-018-0718-3
Bogaard A, Fraser R, Heaton THE et al (2013) Crop manuring and intensive land management by Europe’s first farmers. Proc Natl Acad Scie USA 110:12589–12594. https://doi.org/10.1073/pnas.1305918110
Bohn U, Gollup G, Hettwer C, Neuhäuslová Z, Raus T, Schlüter H, Weber H (2004) Karte der natürlichen Vegetation Europas: Maßstab 1:2 500 000 (map of the natural vegetation of Europe: scale 1:2 500 000). Bundesamt für Naturschutz, Bonn
Boivin N, Crowther A (2021) Mobilizing the past to shape a better anthropocene. Nat Ecol Evol 5:273–284
Bojňanský V, Fargašová A (2007) Atlas of seeds and fruits of Central and East-European Flora: the Carpathian Mountains Region. Springer, Dordrecht
Bronk Ramsey C (2009) Bayesian analysis of radiocarbon dates. Radiocarbon 51:1337–1360
Cappers RTJ, Bekker RM, Jans JEA (2012) Digitale Zadenatlas van Nederland (Digital seed Atlas of the Netherlands). Groningen Archaeological Studies 4, 2nd edn. Barkhuis Publishing, Eelde
Chapman J, Gaydarska B, Hale D (2016) Nebelivka: assembly houses, ditches, and social structure. In: Müller J, Rassmann K, Videiko M (eds) Trypillia mega-sites and european prehistory 4100 – 3400 BCE. Themes in Contemporary Archaeology, vol 2. Routledge, London, pp 117–131
Chapman J, Pashkevych G, Miller D (2020) Plant remains. In: Gadyarska B (ed) Early Urbanism in Europe: the Trypillia Megasites of the Ukrainian Forest-Steppe. De Gruyter, Warsaw, pp 409–412
Chernovol D (2012) Houses of the Tomashevskaya Local Group. In: Menotti F, Korvin-Piotrovskiy AG (eds) The Tripolye Culture Giant-Settlements in Ukraine. Formation, development and decline. Oxbow Books, Oxford, pp 182–209
Dal Corso M, Out WA, Ohlrau R et al (2018) Where are the cereals? Contribution of phytolith analysis to the study of subsistence economy at the Trypillia site Maidanetske (ca. 3900 – 3650 BCE), central Ukraine. J Arid Environ 157:137–148. https://doi.org/10.1016/j.jaridenv.2018.06.009
Dal Corso M, Hamer W, Hofmann R et al (2019) Modelling landscape transformation at the Chalcolithic Tripolye mega-site of Maidanetske (Ukraine): Wood demand and availability. Holocene 29:1622–1636. https://doi.org/10.1177/0959683619857229
Dal Corso M, Pashkevych G, Filipović D et al (2022) Between cereal agriculture and animal husbandry: millet in the early economy of the North Pontic region. J World Prehist 35:3–4. https://doi.org/10.1007/s10963-022-09171-1
Dal Corso M, Kirleis W (in press) Plant economy and local environment at the Trypillia mega-site maidanetske based on botanical macro-remains. In: Hofmann R, Videiko MY, Rud V, Ţerna S, Kirleis W, Müller J (eds) From Ros to Prut: transformations of Trypillia settlements. Sidestone Press, Leiden
Dreibrodt S, Hofmann R, Sipos G et al (2020) Holocene soil erosion in Eastern Europe - land use and/or climate controlled? The example of a catchment at the giant Chalcolithic settlement at Maidanetske, central Ukraine. Geomorphology 367:107302. https://doi.org/10.1016/j.geomorph.2020.107302
Dreibrodt S, Hofmann R, Dal Corso M et al (2022) Earthworms, Darwin and prehistoric agriculture-chernozem genesis reconsidered. Geoderma 409:115607. https://doi.org/10.1016/j.geoderma.2021.115607
Endo E, Nasu H, Haskevych D, Gershkovych Y, Videiko M, Yanevich O (2022) Re-identification of plant impressions on prehistoric pottery from Ukraine. J Archaeol Sci Rep 42:103364. https://doi.org/10.1016/j.jasrep.2022.103364
Ferrio JP, Araus JL, Buxó R, Voltas J, Bort J (2005) Water management practices and climate in ancient agriculture: inferences from the stable isotope composition of archaeobotanical remains. Veget Hist Archaeobot 14:510–517. https://doi.org/10.1007/s00334-005-0062-2
Filatova S (2020) Kakucs-Turján: A case study of macrobotanical formation processes and plant economy from the Hungarian Bronze Age. (Studien zur Archäologie in Ostmitteleuropa 20) Habelt, Bonn
Filipović D, Obradović Đ, Tripković B (2018) Plant storage in neolithic southeast Europe: synthesis of the archaeological and archaeobotanical evidence from Serbia. Veget Hist Archaeobot 27:31–44. https://doi.org/10.1007/s00334-017-0638-7
Filipović D, Meadows J, Dal Corso M et al (2020) New AMS 14C dates track the arrival and spread of broomcorn millet cultivation and agricultural change in prehistoric Europe. Sci Rep 10:13698. https://doi.org/10.1038/s41598-020-70495-z
Filipović D, Kirleis W, Jones G et al (in review) Triticum timopheevii s.l. (’new glume wheat’) in southeastern and eastern Europe: overview of the finds across space and time. Veget Hist Archaeobot
Fischer AD (2021) Urban farmsteads and farming practices. In: Fischer AD, van London H, Blonk-van den Bercken AL, Visser RM, Renes J (eds) Urban farming and ruralisation in the Netherlands (1250–1850). Unravelling farming practice and the use of (open) space by synthesising archaeological reports using text mining. (nederlandse archeologische rapporten). Cultural Heritage Agency of the Netherlands, Amersfoort, pp 115–126
Flohr P, Müldner G, Jenkins E (2011) Carbon stable isotope analysis of cereal remains as a way to reconstruct water availability: preliminary results. Water Hist 3:121–144. https://doi.org/10.1007/s12685-011-0036-5
Fraser RA, Bogaard A, Heaton T et al (2011) Manuring and stable nitrogen isotope ratios in cereals and pulses: towards a new archaeobotanical approach to the inference of land use and dietary practices. J Archaeol Sci 38:2:790–2804. https://doi.org/10.1016/j.jas.2011.06.024
Fraser RA, Bogaard A, Charles M et al (2013) Assessing natural variation and the effects of charring, burial and pre-treatment on the stable carbon and nitrogen isotope values of archaeobotanical cereals and pulses. J Archaeol Sci 40:4:754–4766
Glaser B, Haumaier L, Guggenberger G, Zech W (2001) The ‘Terra Preta’ phenomenon: a model for sustainable agriculture in the humid tropics. Naturwissenschaften 88:37–41. https://doi.org/10.1007/s001140000193
Gluza I (2013) Plant Impressions on the Ceramics. In: Kadrow S (ed) Bilcze Złote: materials of the Tripolye Culture from the Werteba and the Ogród sites, vol 5. Biblioteka Muzeum Archeologicznego w Krakowie, Kraków, pp 87–100
Golea M, García-Vázquez A, Mircea C, Cârciumaru M, Sava G, Mueller J, Kirleis W, Lazăr C (2023) Re-visiting archived rye grains discovered at the neolithic site Cunești (Romania). Radiocarbon 65:1–18. https://doi.org/10.1017/RDC.2023.22
Grikpėdis M, Motuzaite Matuzeviciute G (2018) A review of the earliest evidence of agriculture in Lithuania and the earliest direct AMS date on cereal. Eur J Archaeol 21:264–279
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4. http://palaeo-electronica.org/2001_1/past/issue1_01.htm. Accessed 25 Dec 2022
Heaton THE, Jones G, Halstead P, Tsipropoulos T (2009) Variations in the 13C/12C ratios of modern wheat grain, and implications for interpreting data from bronze age Assiros Toumba, Greece. J Archaeol Sci 36:2:224–2233. https://doi.org/10.1016/j.jas.2009.06.007
Hillman GC (1981) Reconstructing crop husbandry practices from charred remains of crops. In: Mercer R (ed) Farming practice in british prehistory. Edinburgh University Press, Edinburgh, pp 123–162
Hofmann R, Müller J, Shatilo L et al (2019) Governing Tripolye: integrative architecture in Tripolye settlements. PLoS ONE 14:e0222243. https://doi.org/10.1371/journal.pone.0222243
Immel A, Țerna S, Simalcsik A et al (2020) Gene-flow from steppe individuals into Cucuteni-Trypillia associated populations indicates long-standing contacts and gradual admixture. Sci Rep 10:4253. https://doi.org/10.1038/s41598-020-61190-0
Kanstrup M, Thomsen IK, Andersen AJ, Bogaard A, Christensen BT (2011) Abundance of 13C and 15N in emmer, spelt and naked barley grown on differently manured soils: towards a method for identifying past manuring practice. Rapid Commun Mass Spectrom 25:2879–2887. https://doi.org/10.1002/rcm.5176
Kirleis W, Dal Corso M (2016) Trypillian Subsistence Economy: animal and Plant Exploitation. In: Müller J, Rassmann K, Videiko M (eds) Trypillia mega-sites and european prehistory 4100 – 3400 BCE. Themes in Contemporary Archaeology, vol 2. Routledge, London/New York, pp 195–205
Köppen W, Geiger R (1939) Handbuch der Klimatologie, Band 3: Regionale Klimakunde: Europa und Nordasien, Teil N: Klimakunde von Russland in Europa und Asien, 2. Hälfte (Teil N2): Tabellen. Borntraeger, Berlin
Körber-Grohne U (1996) Pflaumen, Kirschpflaumen, Schlehen. Heutige Pflanzen und ihre Geschichte seit der Frühzeit. Theiss, Stuttgart
Kreuz A (2012) Die Vertreibung aus dem Paradies? Archäobiologische Ergebnisse zum Frühneolithikum im westlichen Mitteleuropa. Ber RGK 91/2010:23–196
Kruts VA, Korvin-Piotrovskiy AG, Menotti F, Ryzhov SN, Chernovol DK, Chabanyuk VV (2008) The Trypillia Culture Giant settlement of Talianki. Investigations in 2008. Institute of Archaeology of the National Academy of Sciences of Ukraine, Kyiv
Kruts VA, Korvin-Piotrovskiy AG, Menotti F, Ryzhov SN, Tolochko DV, Chabanyuk VV (2009) The Trypillia Culture Giant settlement of Talianki. Investigations in 2009. Institute of Archaeology of the National Academy of Sciences of Ukraine, Kyiv
Lawrence D, Wilkinson TJ (2015) Hubs and upstarts: pathways to urbanism in the northern Fertile Crescent. Antiquity 89:328–344. https://doi.org/10.15184/aqy.2014.44
Madsen T (2016) CAPCA (Microsoft Excel add-in) Version 3.1. https://www.archaeoinfo.dk/. Accessed 6 Feb 2023
Makarewicz CA, Hofmann R, Videiko MY, Müller J (2022) Community negotiation and pasture partitioning at the Trypillia settlement of Maidanetske. Antiquity 96:831–847. https://doi.org/10.15184/aqy.2022.32
Mathieson I, Alpaslan-Roodenberg S, Posth C et al (2018) The genomic history of southeastern Europe. Nature 555:197–203
Miedaner T, Longin F (2017) Unterschätzte Getreidearten. Einkorn, Emmer, Dinkel & Co, Clenze
Mora-González A, Fernandes R, Contreras Cortés F, Granados-Torres A, Alarcón García E, Delgado-Huertas A (2019) Reporting atmospheric CO2 pressure corrected results of stable carbon isotope analyses of cereals remains from the archaeological site of Peñalosa (SE Iberian Peninsula): agricultural and social implications. Archaeol Anthropol Sci 11:1995–2005. https://doi.org/10.1007/s12520-018-0650-6
Müller J, Pollock S (2016) Trypillia and Uruk. In: Müller J, Rassmann K, Videiko M (eds) Trypillia mega-sites and european prehistory 4100–3400 BCE (Themes in Contemporary Archaeology 2). Routledge, London, pp 281–288
Müller J, Rassmann K (2016) Introduction. In: Müller J, Rassmann K, Videiko M (eds) Trypillia mega-sites and european prehistory 4100–3400 BCE. (Themes in Contemporary Archaeology 2). Routledge, London, pp 1–7
Müller J, Hofmann R, Brandtstätter L, Ohlrau R, Videiko M (2016) Chronology and demography: how many people lived in a mega-site? In: Müller J, Rassmann K, Videiko M (eds) Trypillia mega-sites and european prehistory 4100 – 3400 BCE. (Themes in Contemporary Archaeology 2) Routledge, London, pp 133–170
Müller J, Hofmann R, Kirleis W et al (2017) Maidanetske 2013: New excavations at a Trypillia Mega-site (Studien zur Arch–ologie in Ostmitteleuropa 16). Habelt, Bonn
Müller J, Hofmann R, Ohlrau R, Shatilo M (2018) The social constitution and political organisation of Tripolye mega-sites: hierarchy and balance. In: Meller H, Gronenborn D, Risch R (eds) Überschuss ohne Staat: Politische Formen in der Vorgeschichte (surplus without state: political Forms in Prehistory). Tagungen des Landesmuseums für Vorgeschichte Halle, vol 18. Landesmuseum für Denkmalpflege und Archäologie Sachsen-Anhalt, Halle (Saale, pp 247–262)
Nitsch EK, Charles M, Bogaard A (2015) Calculating a statistically robust δ13C and δ15N offset for charred cereal and pulse seeds. STAR: Sci Technol Archaeol Res 1:1–8
Ohlrau R (2020) Maidanets’ke. Development and decline of a Trypillia mega-site in Central Ukraine. (Scales of Transformation in prehistoric and archaic Societies), vol 7. Sidestone Press, Leiden
Ohlrau R (2022) Trypillia mega-sites: neither urban nor low-density? J Urban Archaeol 5:81–100. https://doi.org/10.1484/J.JUA.5.129844
Pashkevych G (2014) Data of the Trypillian agriculture: new palaeobotanical evidence. In: Dumitroaia G, Preoteasa C, Nicola C-D (eds) Cucuteni culture within the European Neo-Eneolithic context. International Colloquium, Cucuteni 130 Conference Abstracts. Constantín Matasă, Piatra-Neamţ, pp 24–27
Pashkevych GO, Kosakivskyi VA (1991) Rezultaty paleobotanichnoho doslidzhennia trypilskoho poselennia Chechelnyk (The results of paleobotanical investigation of the Trypillia site of Chechelnyk). Tezy dopovidei 10-i Vinnytskoi istoryko-kraieznavchoi konferencii, pp 14–15
Pashkevych GO, Videiko MY (2006) Rilnytsvo plemen trypilskoi kultury (arable farming in the trypillian culture). Naukova Dumka, Kyiv
Philippsen B, Olsen J, Sørensen SA, Måge B (2019) δ13C values of wood and charcoal reveal broad isotopic ranges at the base of the food web. Radiocarbon 61:2:003–2017. https://doi.org/10.1017/RDC.2019.138
Poole I, Braadbaart F, Boon JJ, van Bergen PF (2002) Stable carbon isotope changes during artificial charring of propagules. Org Geochem. https://doi.org/10.1016/S0146-6380(02)00173-0
Pötter S, Schmitz A, Lücke A et al (2021) Middle to late pleistocene environments based on stable organic carbon and nitrogen isotopes of loess-palaeosol sequences from the Carpathian Basin. Boreas 50:184–204. https://doi.org/10.1111/bor.12470
Rassmann K, Ohlrau R, Hofmann R, Mischka C, Burdo N, Videjko MY, Müller J (2014) High precision Tripolye settlement plans, demographic estimations and settlement organization. J Neolithic Archaeol 16:96–134
Reimer PJ, Austin WEN, Bard E et al (2020) The IntCal20 Northern hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62:725–757. https://doi.org/10.1017/RDC.2020.41
Rud V, Hofmann R, Kosakivskyi V, Zaitseva O, Müller J (2019) Trypillia megasites west of the River Southern bug: preliminary results of Bilyi Kamin site investigation in 2018. J Neolithic Archaeol 21:27–60. https://doi.org/10.12766/jna.2019.2
Rud V, Hofmann R, Kosakivskyi V, Zaitseva O, Mueller J (2022) The Pottery Kilns at the Copper and Late Roman age Site Hariachkivka 8 (Ukraine): Magnetic Prospection and Archaeological Verification. In: 16th International Conference Monitoring of Geological Processes and Ecological Condition of the Environment, Nov 2022. European Association of Geoscientists & Engineers, Utrecht, pp 1–5. https://doi.org/10.3997/2214-4609.2022580265
Saile T (2020) On the Bandkeramik to the east of the Vistula River: at the limits of the possible. Quat Int 560–561:208–227. https://doi.org/10.1016/j.quaint.2020.04.036
Schlütz F, Dal Corso M, Kirleis W (in press) The contribution of Chalcolithic terrestrial snail shells from Maidanetske to environmental reconstruction. In: Hofmann R, Videiko MY, Rud V, Ţerna S, Kirleis, W, Müller J (eds) From Ros to Prut: Transformations of Trypillia settlements. Sidestone Press, Leiden
Shatilo L (2021) Tripolye typo-chronology: mega and smaller sites in the Sinyukha river basin. Sidestone Press, Leiden
Styring AK, Carmona CU, Isaakidou V, Karathanou A, Nicholls GK, Sarpaki A, Bogaard A (2022) Urban form and scale shaped the agroecology of early ‘cities’ in northern Mesopotamia, the Aegean and Central Europe. J Agrar Change 22:831–854. https://doi.org/10.1111/joac.12497
Tarongi M, Prats G, Alonso N (2020) The storage of pulses during the bronze and Iron Ages in the East of the Iberian Peninsula: examining the archaeological data through the lens of ethnography. J Archaeol Sci Rep 30:102174. https://doi.org/10.1016/j.jasrep.2019.102174
Terna S (2021) How can we investigate the socio-economic structure of the Cucuteni-Tripolye mega-sites? Houses as snapshots and pits as durable narratives. Arch Korrbl 51:37–60
Treasure ER, Church MJ, Gröcke DR (2016) The influence of manuring on stable isotopes (δ13C and δ15N) in Celtic bean (Vicia faba L.): archaeobotanical and palaeodietary implications. Archaeol Anthropol Sci 8:555–562. https://doi.org/10.1007/s12520-015-0243-6
Vaiglova P, Snoeck C, Nitsch E, Bogaard A, Lee-Thorp J (2014) Impact of contamination and pre-treatment on stable carbon and nitrogen isotopic composition of charred plant remains. Rapid Commun Mass Spectrom 28(2):497–2510. https://doi.org/10.1002/rcm.7044
Van der Veen M, Jones G (2006) A re-analysis of agricultural production and consumption: implications for understanding the British Iron Age. Veget Hist Archaeobot 15:217–228. https://doi.org/10.1007/s00334-006-0040-3
Videiko M (2017) Search for plant prints on a burnt daub from Maidanetske. https://www.academia.edu/36738285/_Archaeology_Mariia_Videiko_Search_for_plant_prints_on_a_burnt_daub_from_Maidanetske. Accessed 21 Feb 2023
Wallace M, Jones G, Charles M, Fraser R, Halstead P, Heaton THE, Bogaard A (2013) Stable carbon isotope analysis as a direct means of inferring crop water status and water management practices. World Archaeol 45:388–409. https://doi.org/10.1080/00438243.2013.821671
Wiedner K, Schneeweiß J, Dippold MA, Glaser B (2015) Anthropogenic Dark Earth in Northern Germany—The Nordic Analogue to terra preta de Índio in Amazonia. CATENA 132:114–125. https://doi.org/10.1016/j.catena.2014.10.024
Yanushevich ZV (1975) Fossil remains of cultivated plants in the South-West of the Soviet Union. Folia Quat 46:23–30
Yanushevich ZV (1978) Prehistoric food plants in the south-west of the soviet-union. In: Behre K-E, Lorenzen H, Willerding U (eds) Beiträge zur Paläo-Ethnobotanik von Europa (contributions to the palaeo-ethnobotany of Europe). Berichte der Deutschen Botanischen Gesellschaft 91, Special Issue. Fischer, Stuttgart, pp 59–66
Yanushevich ZV (1980) Zemledelie v rannem tripoije (podannym palaeobotaniki). In: Artemenko II (ed) Pervobytnaya archeologiya: poiski I nakhodki. Naukova Dumka, Kyiv, pp 225–234
Zohary D, Hopf M, Weiss E (2012) Domestication of plants in the Old World, 4th edn. Oxford University Press, Oxford
Acknowledgements
We thank Svetlana Ibens for help in the field; Tanja Reiser for support with the macrobotanical analyses; Dragana Filipović for cross-checking the identification of the charred Timopheev’s wheat remains; Esther Thelen, Carsten Reckweg and Nicole Taylor for producing and improving the figures; students Pia Bodden and Ricarda Liedtke at Kiel University and Nikita Kuzemko from Uman, as well as other students and colleagues, for helping with collecting samples and carrying out flotation; Pascal Flohr and Cheryl Makarewicz at Kiel University for initial assistance with Fourier-transform infrared spectroscopy and stable isotope analyses; Steven Bouillon and Yannick Stroobandt at the stable isotope laboratory of the Earth and Environmental Science Department of the Katholieke Universiteit at Leuven for processing the stable isotope samples, Suzanne Needs-Howarth, Toronto, for copy editing and two anonymous reviewers for insightful comments and high appreciation. We also thank I.G. Emelyanov, director of the National Museum of Natural History of the National Academy of Sciences of Ukraine at Kyiv, for archiving the charred plant remains recovered by Galyna Pashkevych. We are grateful for funding provided by the Deutsche Forschungsgemeinschaft (German Research Foundation) within the programme of the Collaborative Research Centre (CRC) 1266 Scales of Transformations in Prehistoric and Archaic Societies [Project Number 2901391021–Sonderforschungsbereich (SFB) 1266].
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by F. Bittmann.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kirleis, W., Corso, M.D., Pashkevych, G. et al. A complex subsistence regime revealed for Cucuteni–Trypillia sites in Chalcolithic eastern Europe based on new and old macrobotanical data. Veget Hist Archaeobot 33, 75–90 (2024). https://doi.org/10.1007/s00334-023-00936-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-023-00936-y