
Abstract
The archaeological excavation of two abandoned prehispanic agricultural terraces (Infiernillo and Tocotoccasa) in the Chicha-Soras Valley (Apurimac) in southern-central Peru revealed the presence of palaeosols. The palaeosols represent soil that developed following construction of agricultural terraces during the Middle Horizon. The soil profiles at the current surface developed following reconstruction of the terraces during the Late Intermediate Period. Phytolith analysis revealed an unexpected presence of Arecaceae (palm family) and Marantaceae (arrowroot family) in both terraces, which has been attributed to local cultivation and/or transportation and use of soil, dung, plant material or implements (made of Arecaceae) on the terrace surfaces. Pollen analysis of a nearby wetland (Ayapampa) did not provide evidence for Arecaceae or Marantaceae. Both phytolith and pollen analysis of the terraces and wetland (respectively) indicated that Zea mays was cultivated locally during the Middle Horizon and Late Intermediate Period, although phytoliths of maize are absent from the wetland record during the Middle Horizon. The presence of Solanaceae and Chenopodiaceae/Amaranthaceae pollen in the wetland may be indicative of cultivation of further important taxa during the Middle Horizon and Late Intermediate Period, which continued into the Late Horizon together with Zea mays.
Similar content being viewed by others


Introduction
Terrace agriculture was an important feature of the economy of the Peruvian Andes during Pre-Hispanic times. The terraces, which blanket much of the landscape, take many different forms (e.g. cross channel, sloping field, linear bench and irrigated bench), and their construction is believed to be strongly related to their function, elevation, local geomorphology, regional precipitation and temperature patterns, and crop selection practices (Osgood Brooks 1998). In regions where archaeological studies have been conducted, it has been hypothesised that terrace construction and morphological change represent a developmental history that may have been a necessity for increasing productivity and yield due to socio-economic pressures, possibly linked to climate variability and environmental change (e.g. Osgood Brooks 1998; Kemp et al. 2006; Branch et al. 2007; Goodman-Elgar 2008). Whatever the forcing factor, or factors, they represent an extraordinary technological achievement and modification of the natural environment that was undoubtedly an adaptation strategy to increase resilience and reduce risk.
The preservation of sub-fossil biological remains in ancient soil sequences has the potential to provide significant information on the history of crop selection, biomass burning, and irrigation practices. Such studies, when conducted on terraces with deposits spanning their full developmental history, would enable reconstruction of the changes of these practices through time. Such data, combined with archaeological records, and a secure chronological framework, and ideally palaeoecological records from adjacent wetlands, would enable important research questions of relevance to Central Andean land use systems as a whole to be addressed. These include: What was the precise timing of terrace use and the nature of cultivation? Is there evidence for multicropping and changes in crop selection? Is there evidence for deliberate burning and irrigation of terrace surfaces? Is there a clear relationship between the timing of terrace use (re-use) and abandonment, and periods of wider socio-economic change, and/or climate variability and environmental change? Is there evidence for the transportation and utilisation, and possibly the cultivation, of plant taxa outside the immediate ecozone? Given the importance of these questions, it is surprising that no detailed archaeobotanical studies, especially phytolith analysis, have been conducted on pre-Hispanic terraces especially considering their role in Andean agricultural systems. Instead, studies have focused on a range of archaeological sites and agricultural features located in both the coastal and highlands of Peru, including mounds (Grobman et al. 2012; Dillehay 2017), ceremonial sites (Tykot et al. 2006; Sayre 2010; Castillo Luján 2019), human remains (Correa Trigoso 2018), settlements (Lavallée et al. 1999, 2011; Perry et al. 2006, 2007; Chevalier 2008; Caramancia et al. 2018) and corrals (Duncan 2003). Whilst these studies have provided valuable information on crop selection and the environmental context of human activities through phytolith analyses, the potential of phytolith studies on agricultural terraces has not been explored.
Phytoliths analysis can provide information of the past use of agricultural terraces with phytolith assemblages largely reflecting on-site plant usage. Phytoliths are opaline silica bodies which form in the leaves, seeds, fruits, reproductive systems, and roots of plants, and are released into the soil when a plant dies and decomposes. For this reason, they are largely considered an in-situ deposit (Piperno 2001, 2006; Watling and Iriarte 2013). Phytoliths may, however, be added to an assemblage via slope wash processes and human disturbance in the landscape (e.g. land clearance for agriculture) as well as being liberated into the air during fire events (Piperno 2001, 2006). Phytoliths of certain cultivars, such as maize, are highly diagnostic as they produce different morphotypes within their leaves, stalks, and cobs. In particular, wavy-topped rondels produced in the cobs and fruitcases of maize are diagnostic of cultivated maize (Ball et al. 2016). Maize is considered to be a very important crop within the Peruvian highlands, with an increase in terracing during the Middle Horizon (~ 1,350–950 cal bp; ad ~ 600–1000) being linked to the expansion of maize cultivation (Schreiber 1992; Cook 2004). The construction of pre-Hispanic terraces increased the area of land suitable for cultivation whilst controlling erosion, deepening the soil matrix and possibly creating microclimates aiding frost prevention, and regulating soil temperature and humidity (Lane 2014).
Archaeological context
The first indication of occupation with some permanence in the Chicha-Soras Valley was during the Initial Period (~ 3,750 − 2,850 cal bp; ~1800 − 900 bc). There are very limited signs for human activity in the area from the late Initial Period until the Middle Horizon (~ 1,350–950 cal bp; ad ~ 600–1000) with two small sites having been located for this interval, indicative of a low-density human presence over this period. The adjoining Sondondo Valley ~ 40 km to the west and the Andahuaylas area to the northeast have both provided remains of Early Horizon (~ 2,850 − 2,150 cal bp; ~900 − 200 bc) and Early Intermediate Period (~ 2,150 − 1,350 cal bp; ~200 bc-ad 600) occupation. For the Chicha Soras valley 13 sites with Middle Horizon activity have been identified; of these, there are 7 with evidence of occupation, all of which were situated south of the present-day village of San Pedro de Larcay. Middle Horizon sites include Yako (3,330 m a.s.l.), which has surface and excavated Ocros, Viñaque and Huamanga style ceramics, and chert, basalt and obsidian lithics, as well as copper alloy objects and metal working slag. Architectural remains found on this 0.65 ha site include two large circular structures measuring 7 − 8.5 m in diameter as well as at least one rectilinear structure and a single D shaped building. Further Middle Horizon sites on the west bank of the Rio Chicha include Chiqna Jota, situated 450 m south of the confluence of the Rio Pachachaca with the Rio Chicha. At 3,460 m a.s.l., parts of the architectural core of Chiqna Jota include a gallery type structure on a north south alignment on the east side of the plaza, and ~ 30 m further north are the remains of a rectilinear structure that would originally have had multiple stories. The Middle Horizon witnessed the introduction of extensive irrigated agricultural terracing (Keeley and Meddens 1993; Kemp et al. 2006; Branch et al. 2007; Meddens and Branch 2010), with four types of terraces having been identified (Kendall and Rodriguez 2009). In the upper reaches of the Yanomayo drainage, a tributary to the Rio Chicha the Naupallacta site complex is situated which served to manage the camelid herds in the area and which was supported by the agricultural sites in the Chicha Soras valley. The end of the Middle Horizon, with the collapse of the Wari polity, saw some sites in the Chicha-Soras Valley being permanently abandoned, while others continued being occupied.
The Late Intermediate Period (~ 950 − 474 cal bp; ad ~ 1000–1476) in the Chicha-Soras Valley, and on its adjoining altiplano, witnessed significant population growth, with the number of occupation sites increasing from 7 to 22. Not only did the number of settlement sites increase but also their size e.g. Laymi (3,360 m a.s.l.), measuring ~ 25 ha; Taccarampa (3,570 m a.s.l.) covering ~ 20–25 ha. Chiqna Jota expands to ~ 9 ha around this time with up to 200 mostly circular structures arranged in small groups around communal patios. This important valley wide population growth accompanied the emergence of defensive structures at sites, such as the fortification of the hill on the eastern margin of Chiqna Jota. Qasamarca/Qasapampa similarly had a mountaintop with defensive walling rising ~ 50 m above the lowermost level. A further demographic shift takes place in the late, Late Intermediate Period (ad ~ 1200–1476) from valley-based sites to settlements such as Puyca, Auquimarca, Chuntaya/Jasinchilla, Qasinchilla, Uchuy Umay and Ccanchu sited on mountain peaks on the altiplano, often accompanied by perimeter walling along the sides, which afforded easiest access to the summits. The shift of settlements to defendable topographical settings and the appearance of fortifications was a widespread and common occurrence in the central and southern Andes (Arkush 2011). The differentiation between the early and late, Late Intermediate Period has also been recognised in the pottery sequence for the Chicha-Soras Valley. The earlier part being characterised by Chicha/Chanca style assemblages, with similarities to and equivalents in Tanta Orqo and Toqsa style ceramics (Meddens and Vivanco 2018). The late Late Intermediate Period assemblages largely comprise Soras and Arqalle style pottery with the Soras material having concurrences with the Aya Orqo style materials (Meddens and Vivanco 2018) as found in Ayacucho and parts of Apurimac and Huancavelica.
During the late Late Intermediate Period there is evidence for extensive remodelling, reuse and expansion of the earlier terraced agricultural systems associated with the further construction of large-scale irrigation management systems (Kemp et al. 2006; Branch et al. 2007; Kendall and Rodriguez 2009). There are also widespread Late Intermediate Period occupation sites directly associated with the agricultural terracing and irrigation infrastructure, such as Taccarampa, Apuraccay, Cupo, Aputacca, Montaccahua on the west and Kulkunchapampa and Huaychuapata on the east bank of the river, as well as the previously mentioned defensive sites of Puyca, Auquimarca, Chuntaya/Jasinchilla, and Qasinchilla. For the Late Horizon (~ 474 − 416 cal bp; ad ~ 1476–1534), there was a consolidation in the number of habitation sites from 22 for the preceding Late Intermediate Period to 18, with a reduction in the high-altitude mountain top sites, and expansion of the size and number of mid-valley, slope settlements. Iglesiachayoc was established at 3,410 m a.s.l., as was Soras at 3,425 m a.s.l. at the north end of the valley. At the same time, Chiqna Jota, Qasamarca, Laymi and Taccarampa get significant provincial Inca architectural elements added at to the central sectors of these sites.
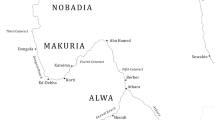
Here we present the findings from a detailed phytolith study of pre-Hispanic agricultural terraces in the Chicha-Soras Valley (Apurimac). Long-term research in the valley by Meddens (e.g. Meddens and Vivanco 2018) led to the initiation of a fully integrated archaeological and palaeoenvironmental study of ancient agricultural terraces and their associated wetlands. A detailed account of the palaeopedological and geochemical analysis of two of the terrace profiles, Infiernillo (INF) (14°12’29.4”S, 73°32’25.1”W; 3,447 m a.s.l.) and Tocotoccasa (TOC) (14°11’40.1”S, 73°32’18.9”W; 3,417 m a.s.l.), was reported in Kemp et al. (2006) and Branch et al. (2007) (Fig. 1). These studies revealed the presence of multi-period human occupation and a number of buried palaeosols within terrace sections in close proximity to a wetland (Ayapampa). The main characteristics of the macro- and micromorphology of INF (ESM Table S2) revealed the presence of a clearly defined 2bAh horizon overlying a strongly weathered 2bBt horizon collectively representing a buried terrace soil profile. Radiocarbon dates from three different pieces of wood charcoal in the 2bAh (ESM Table S1) indicate that the terrace is Middle Horizon in age which is also confirmed by the presence of Middle Horizon pottery. The main characteristics of the macro- and micromorphology of TOC (ESM Table S3) revealed the presence of two buried soils: the original sloping land surface (3bAh), and the surface of a former agricultural terrace (2bAh). A radiocarbon date from a single piece of wood charcoal in the 2bAh (ESM Table S1) indicates that the terrace is Middle Horizon in age. Late Intermediate Period pottery on the present-day terrace surface (Ah and Bw horizons) indicates the possible timing of terrace reconstruction. The total phosphate and plant-available phosphate are higher in the Ah and 2bAh horizons, which were interpreted as evidence for the effect of manuring (Branch et al. 2007). Therefore, the developmental history of both terraces, together with the adjacent palaeoecological record from the wetland, provided an excellent opportunity to address the research questions outlined earlier.

Location of the Chicha-Soras Valley, Infiernillo (INF) and Tocotoccasa (TOC) agricultural terraces, Ayapampa wetland and archaeological sites mentioned in the text. Figure produced using ArcGIS 10.5.1
Methods
The archaeological excavation and sampling of the ancient terraces involved cutting a trench (~ 1 × 10 m) perpendicular to the terrace wall, description of the soil profile within a 1 m wide section (Hodgson 1976), and the recovery of undisturbed blocks (7 × 5 × 4 cm) for thin section preparation and micromorphological analysis (Kemp et al. 2006). Furthermore, extraction of bulk samples at 5 cm intervals from the section permitted analysis of total organic carbon (%) and total and available phosphate (Branch et al. 2007), and phytoliths. Core samples, obtained from the Ayapampa wetland using a Russian peat sampler (semi-circular, 5 cm diameter), were described using the Troels-Smith method (1955), and analysed for organic matter content, pollen and microscopic charred particles (Branch et al. 2007), and an assessment of the phytolith content (Handley 2022).
In order to extract the phytoliths, each sample was sieved through a 500 μm sieve and 5 g dry weight of material < 500 μm was dispersed with Hydrochloric acid to remove carbonates. Sodium hexametaphosphate was then added to remove the clay fraction. Samples were transferred to a furnace and heated at 500 °C for 2.5 h to remove organic matter. Sodium polytungstate (specific gravity of 2.3 g/cm3) was used to separate the remaining mineral and fine organic fraction from the phytolith concentration, quartz however cannot be separated from the phytoliths as they have the same density; quartz is easily identifiable and distinguishable from the phytoliths as it is visible under cross polarised light unlike phytoliths. Finally, the samples were dehydrated using either a lamp (Tocotoccasa - TOC) or ethanol (Infiernillo - INF) before being mounted on microscope slides in Entellan New™. Samples were processed in a laboratory not used for processing of lowland tropical phytoliths to avoid potential contamination with extra-regional phytolith morphotypes. A total of 300 phytoliths (all morphotypes) were counted per slide; raw counts are provided in ESM Tables S4 and S6. Identification was aided by published literature on phytoliths forms especially Piperno (2006), as well as online databases including PhytCore (https://www.phytcore.org/phytolith/index.php), and the Tropical Palaeoecology Phytolith reference collection housed at the University of Reading. Phytolith nomenclature follows the International Code for Phytolith Nomenclature 2.0 (ICPT 2019). A number of phytoliths diagnostic of Poaceae were encountered, including grass silica short-cell phytoliths (GSSCP) produced in grass epidermal cells; Trapezoid, Saddle, Polylobate, Bilobate, Cross, Crenate and Rondel phytoliths. Trapezoid and Crenate phytoliths are commonly found in the Pooideae sub-family, while Polylobate and Bilobate are diagnostic of Panicoideae sub-family grasses, and Saddles of Chloridoideae, allowing for the identification of grass sub-families within the phytolith assemblage. Acute bulbosus phytoliths are thought to be diagnostic of Poaceae, along with Elongate sinuate phytoliths, especially when found in association with each other (ICPT 2019). Phytoliths diagnostic of Cyperaceae include: Cyperaceae conical phytoliths, Cyperaceae hat-shaped, and Cyperaceae achene epidermal phytoliths (Piperno 2006). Bulliform flabellate phytoliths are produced in both Poaceae and Cyperaceae, and a high number of these phytoliths can be produced where water availability is high (Fisher et al. 2013). Elongate entire phytoliths have low diagnostic value as they are found in grasses, monocot, eudicots and conifers (Strömberg 2003). Tracheary phytoliths are commonly found in arboreal plants (Piperno 2006; Dickau et al. 2013).
Additional bulk sub-samples from the terrace sections were sorted with clean fine forceps and fragments of charcoal isolated for radiocarbon dating (Branch et al. 2007). Although minute amounts of wood charcoal were present in most samples, only the Ah and bAh horizons contained significant quantities. Samples of wood charcoal within each of the palaeosol Ah horizons were submitted to the NERC Radiocarbon Laboratory East Kilbride for AMS dating. Radiocarbon ages for the terraces were calibrated using OxCal v4.4 and the southern hemisphere calibration curve (SHCal 20), and are presented as individual dates in ESM Table S1. For the Ayapampa wetland sequence, the chronology is provided through Accelerator Mass Spectrometry (AMS) radiocarbon measurements on bulk peat samples, taken from the core at lithostratigraphic and biostratigraphic unit boundaries. Six measurements (Wk-samples) were submitted to the Waikato Radiocarbon Dating Laboratory, New Zealand and one sample (Beta-142,326) was analysed by Beta Analytic Inc., USA. Details of the dated samples, radiocarbon ages, and associated stable isotopic measurements are provided in ESM Table S1. The radiocarbon results are conventional radiocarbon ages (Stuiver and Polach 1977). The age-depth model (ESM Fig. S1) has been constructed using the program OxCal v4.2 (Bronk Ramsey 2009; Bronk Ramsey and Lee 2013) and the southern hemisphere (SHCal 13) atmospheric calibration curve (Hogg et al. 2013). The P_Sequence Poisson process model (Bronk Ramsey 2008) employs a variable k parameter (Bronk Ramsey and Lee 2013) with the overall age-depth model defined as P_Sequence (1,1,U(-2,2), with k0 (the base k parameter) = 1 cm− 1, the interpolation rate = 1 cm− 1 (output from the model given every 1 cm), and variability in k allowed between a factor of 10− 2 and 102. The surface of the basin was modelled as the sampling date (ad 2001 ± 5 years). Even with the use of a P-sequence model, it was not possible to create a stable age-depth model that took account of all dates. Examination of the dates, stratigraphy, and the pollen record highlighted two potential hiatuses, related to minerogenic sediment deposition. Subsequent use of two separate P-sequence models, and no modelling of the basal date allowed for a stable overall model to be generated. The overall sequence provided an Amodel value of 98 and was considered to be a good fit to the data, which can be seen in the plot (ESM Fig. S1).
Results
Phytolith analysis of the ancient agricultural terraces
The high abundance of phytoliths of the Pooideae sub-family (Poaceae) throughout the INF sequence indicates C3 grasses formed the dominant vegetation community (Fig. 2; ESM Table S5). There is also a signal for C4 plants, with high proportions of phytoliths of the Chloridoideae sub-family (Poaceae) and a continuous signal of the Panicoideae sub-family (Poaceae) throughout. The latter includes Zea mays (maize), which has been cultivated in highland Peru for at least 4,000 years (Perry et al. 2006). This is confirmed by Wavy top Rondel phytoliths present throughout the soil sequence, which indicate that maize was grown on the terraces (see Piperno 2009). Maize is thought to have originated in the Central Balsas River Valley of Mexico by 8,700 cal bp (based on the analysis of phytoliths and starch grains), and spread into Colombia by 7,000–6,000 cal bp, with some of the earliest dates for its occurrence in Peru being 3,600-4,000 cal bp. In the Zaña Valley in northern Peru, early cultivation of beans, squash and coca was accompanied by irrigation (Dillehay et al. 2005), attesting to the body of evidence for widespread food production by 6,000 cal bp. The presence of large Type 1 Cross phytoliths, as found in this study, can also be an indicator of cultivated maize because they occur in the leaves of maize plants. Further analysis would be required, however, using discriminant function analysis (see Piperno 2006, p 49), to determine whether the Cross-shaped phytoliths within these samples were cultivated maize or other wild Panicoideae sub-family grasses. The presence of Acute bulbosus, Elongate sinuate, and Rondel phytoliths confirms the dominance of grasses on the terrace surface and probably within the general environment. The presence of a large number of grass phytoliths, despite the agricultural use of these terraces, could be due to several factors including the presence of weeds alongside cultivated crops, the regrowth of grasses during periods of abandonment and grasses growing on the edges of the terraces and the adjacent slopes. They may have also been incorporated in the soil profile via the use of camelid dung as a fertiliser, with the camelids likely grazing on the Ayapampa wetland which is known to have been colonised by Poaceae (see section below). Sedge (Cyperaceae) phytoliths, including Cyperaceae achene and Cyperaceae hat shaped phytoliths indicate wet soils either on the terrace surface or near to the terraces, possibly adjacent to the irrigation canals. This interpretation is supported by the presence of Bulliform flabellate phytoliths, which are indicative of wetland grasses; the production of Bulliform flabellate phytoliths is therefore likely a consequence of an excess of water within the growing environment promoting the growth of aquatic grasses (Piperno 2006, p 35).

Phytolith diagram for Infiernillo (INF) for all encountered phytolith morphotypes. The phytolith diagram was produced using TILIA and TILIA*GRAPH (Grimm 1991–1993) and the soil profile is modified from Kemp et al. 2006. INF 70 (1, 2 and 3) indicates the sampling location for radiocarbon dating, for more details on the dating see ESM Table S1
The phytolith composition of TOC is similar to that at INF with high levels of Pooideae sub-family phytoliths throughout the profile indicating C3 grasses (Fig. 3; ESM Tables S6, S7). There is a lower proportion of Chloridoideae sub-family Saddle phytoliths whilst sedge phytoliths occur in similar proportions to INF indicating wet soils on the terrace or nearby. There is a continuous signal of C4 plants with Panicoideae sub-family phytoliths throughout. Acute bulbosus, Elongate sinuate and Rondel phytoliths support the interpretation of grass dominance. There is clear evidence for cultivation with maize phytoliths present throughout the soil profile. High levels of burnt phytoliths were found in association with the original land surface (3bAh) identified by the presence of occluded carbon within the phytoliths and a dark opaque colouration (see Parr 2006; Dong et al. 2022), which may indicate burning in order to clear land for the development of the agricultural terraces. The presence of burnt phytoliths were also recorded within the Middle Horizon terrace (2bAh), suggesting the use of fire as part of the farming practices during this time.

Phytolith diagram for Tocctoccasa (TOC) for all encountered phytolith morphotypes. The phytolith diagram was produced using TILIA and TILIA*GRAPH (Grimm 1991–1993) and the soil profile is modified from Branch et al. 2007. TOC 70 indicates the sampling location for radiocarbon dating of the terrace sequence, for more details on the dating see ESM Table S1
The most unexpected signal to come out of analysis of both terrace sequences was the presence of two types of spherical phytoliths. The first being Spheroid echinate phytoliths (Fig. 4a) that almost exclusively belong to species within the Arecaceae (palm) family. Within Peru, palms typically grow in warm, moist montane forests (‘cloud forest’) on the eastern side of the Peruvian Andes from < 3,500 m a.s.l. (Henderson et al. 1995; Galeano et al. 2008). Today, 17 species of palm are strictly Andean, whilst the remainder (123 species) are from lower elevations including Andean/Subandean, strictly Subandean, Amazonia and peripheral species of the Madre de Dios (Kahn and Moussa 1994). The presence of one or possibly more species of this family, as will be discussed further below, in the Chicha-Soras Valley is therefore surprising and suggests contact with the eastern Andes. Evidence for trade between Andean and Amazonian communities has a long history with communities sourcing crops and food, and wood for tools and weapons, unable to be grown within their immediate environment (see Kimura 1985; Suarez and George 2011).

Microphotographs of phytoliths of a Arecaceae (Spheroid echinate) and b Marantaceae (Spheroid ornate)
Palaeoecological investigation of the wetland
The pollen-stratigraphic record for the Ayapampa wetland is reproduced in Fig. 5 and ESM Table S8 (Branch et al. 2007). Here we present an updated interpretation with a new age model. The record indicates that prior to ~ 2,064 cal bp (2,191 − 1,936 cal bp, 315 cm; LPAZ 1 and 2) the landscape surrounding the wetland was dominated by species of grasses (Poaceae), and perennials and shrubs of the daisy family (Asteroideae/Cardueae). The wetland was mainly composed of sedges (Cyperaceae) and probably grasses with herbaceous taxa including Plantago (e.g. P. tubulosa, P. rigida) and alder woodland (Alnus). It is possible that Plantago was one of the main peat forming plants, as it is in many parts of Peru today, alongside Distichia muscoides (Juncaceae) (Salvador et al. 2014/2015). The organic matter values during LPAZ 2 (> 60%), and also parts of LPAZ 4 and 5, suggest that cushion plants together with sedges probably dominated the wetland surface forming a minerotrophic peatland. The decline of sedge pollen values from before ~ 3,628 to ~ 2,284 cal bp (3,909 − 3,455 to 2,534 − 2,034 cal bp) during a period of mineral sediment deposition in the wetland, and re-expansion from ~ 2,284 cal bp coincident with a renewal of peat formation (LPAZ 2), suggests that sedges are a sensitive indicator of hydrological changes. The results indicate that sedge populations declined during periods of higher water level inferred by mineral-rich fluvial sediment from 406 − 375 cm and subsequent rise of pondweed during LPAZ 2 (Potamogeton), which prefers standing or slow flowing freshwater (~ 50 cm in depth). The high proportion of Solanaceae (nightshade family) and Chenopodiaceae/Amaranthaceae (amaranth family) at the time of landscape instability is particularly interesting. Both of these families contain important cultivated plants, including potato (Solanum tuberosum) and quinoa (Chenopodium quinoa), as well as other annual and perennial herbs, and a few shrubs and trees. Archaeological evidence indicates that potato and quinoa have been cultivated in Peru for at least 4,000 years (Harris and Hillman 1989), and their presence in the Chicha-Soras Valley, especially during the early part of LPAZ 2, may therefore have cultural and economic significance for human occupation during the Initial Period (~ 3,750 − 2,850 cal bp; ~1800 − 900 bc) and Early Horizon (~ 2,850 − 2,150 cal bp; ~900 − 200 bc).

(modified from Branch et al. 2007). For each sample, maximum pollen counts of 300 pollen grains and spores were attempted. Microscopic charred particles were counted during the pollen analysis and are presented on the pollen diagram as a % of total pollen
Pollen diagram of selected taxa (% total pollen including aquatics and spores) from the Ayapampa wetland.
LPAZ 3 is marked by declining organic matter values from ~ 2,014 cal bp (2,153 − 1,875 cal bp) to ~ 1,883 cal bp (2,147 − 1,618 cal bp) (300 − 273 cm) that culminate in the deposition of a thick unit of sandy clay (273 − 242 cm). The renewal of peat formation at ~ 692 cal bp (796 − 588 cal bp) means that this significant period of landscape erosion occurred during the later Early Intermediate Period (~ 2,150 − 1,350 cal bp; ~200 bc-ad 600), Middle Horizon (~ 1,350–950 cal bp; ad ~ 600–1000) and into the early part of the Late Intermediate Period (~ 950 − 474 cal bp; ad ~ 1000–1476) cultural periods. Calculation of an age model for this part of the sequence has proven difficult because of these marked sedimentary changes. Nevertheless, once again, higher water levels are indicated by the reduction in sedge and presence of pondweed indicating a major hydrological change in the wetland. Solanaceae, Chenopodiaceae/Amaranthaceae and Zea mays (maize) were all present, signifying a prolonged period of cultivation. The microscopic charred particle data for LPAZ 3 indicates an overall reduction in burning that appears to contradict the pollen evidence for human activity. Burning is frequently used today in many parts of the Peruvian Andes to clear stubble and to aid replenishment of soil nutrients. Therefore, it might signify a reduction in the human activity over time, or simply a change in burning practices.
The lower part of LPAZ 4 is marked by a renewal of peat formation from ~ 692 cal bp (796 − 588 cal bp) and a concomitant increase in a grass dominated wetland. The surrounding landscape continued to be dominated by species of the daisy family (Asteroideae/Cardueae) as well as grasses (Poaceae). The presence of Chenopodiaceae/Amaranthaceae pollen may indicate cultivation during the Late Intermediate Period (~ 950 − 474 cal bp; ad ~ 1000–1476). This interpretation is supported by the presence of maize phytoliths (Wavy top Rondels) from ~ 595 − 512 cal bp i.e. throughout the late, Late Intermediate Period in the wetland (Handley 2022). The upper part of LPAZ 4 in contrast is marked by the deposition of organic lake sediment and an increased influx of mineral rich sediment into the wetland from ~ 611 − 553 cal bp (~ 666 − 556 to ~ 646 − 460 cal bp). The change in hydrology is supported by the presence of pondweed. Solanaceae, Chenopodiaceae/Amaranthaceae and Zea mays were all present suggesting a continuation of farming practices.
The Late Horizon (~ 474 − 416 cal bp; ad ~ 1476–1534) is poorly constrained by the pollen record (~ 118–158 cm; LPAZ 5) but is characterised by possible evidence for cultivation of Solanaceae, Chenopodiaceae/Amaranthaceae and Zea mays, as well as overwhelming evidence for vegetation burning. This interpretation is supported by the presence of maize phytoliths (Wavy top Rondels) at ~ 494 cal bp in the wetland (Handley 2022). It marks a notable transition to a prolonged period during the Colonial Period (~ 416 − 124 cal bp; ad ~ 1534–1826) and Republican Period (ad 1826 to the present day) differentiated by a reduction in burning, increased landscape erosion (except in the last ~ 50 years) and a demise in the frequency of cultivars. It is tempting to suggest that this reflects the decline of large-scale cultivation on agricultural terraces and reduced management of the landscape, including the maintenance of terraces. A change in hydrology of the wetland in the last 50 years or so is indicated by the rise in sedges and reduction in erosion together with the only evidence for cultivation of European cereals.
Discussion
Pre-hispanic terrace agricultural practices in the southern-central Peruvian Andes
The phytolith analysis of the TOC and INF agricultural terraces has demonstrated the clear potential of this method within an Andean context for reconstructing vegetation cover (grasses and sedges) and land-use (maize cultivation). Grassland was the dominant vegetation community indicated by an abundance of C3 Pooideae sub-family phytoliths. This is a common community at high altitude today in Peru and is often in environments with higher moisture availability and lower temperatures (Aleman et al. 2014). The moisture levels are probably a reflection of irrigation systems associated with the terraces creating localised wet pasture/meadow, as well as the proximity of the Ayapampa wetland. Given that Chenopodiaceae/Amaranthaceae taxa do not produce diagnostic phytoliths, and appear silent in the phytolith record, the main indicator of cultivation in the phytolith assemblage is maize. As a result, there is no evidence within this new analysis for multicropping practices on the terraces. Although Chenopodiaceae/Amaranthaceae and Solanaceae were recorded within the pollen diagram (Fig. 5) from Ayapampa, there is no way of confirming whether they were planted alongside maize on the terraces or if they formed an alternative crop selection to maize during certain periods. Maize was present at relatively high abundance within the TOC 2bAh and 2bBt horizons indicating its growth on the Middle Horizon terraces. Its presence in the Ah2 and AB horizons may also indicate maize cultivation during the Late Intermediate Period. At INF, the presence of maize phytoliths, within the 2bAh and 2bABt also suggests Middle Horizon cultivation. Interestingly, more phytoliths from maize cobs (Zea mays Wavy top Rondels) were present within both profiles than those formed within the leaves and husks of the maize plant (Crosses). Maize cobs are normally removed from terraces during harvest resulting in a greater signal of phytoliths from the leaves and stalks of the plant (Watling et al. 2015; Dickau et al. 2016). It is possible that not all maize cobs were removed from the terraces during harvest, or that some of the cobs were returned to the fields after the kernels had been removed as an additional fertilizer. The number of Wavy top Rondels in this study is also surprising when compared to other studies in South America (e.g. Iriarte et al. 2010; Watling et al. 2015; Lombardo et al. 2020) where they were found only in trace amounts (> 1%). Whatever the agricultural techniques employed within the Chicha-Soras Valley there appears to be a greater occurrence of maize cobs on the terrace than one might expect. Further phytolith analysis of terrace deposits from the Andean highlands would provide greater clarity on agricultural practices associated with maize growth at altitude.
One thing we do know, the growth of maize at high altitude in relatively cold and dry Andean environments was certainly enhanced by extensive modification of the landscape with the construction of agricultural terraces creating an appropriate habitat (e.g. soil temperature, moisture, structure, and nutrients). However, maize is known to be susceptible to drought because of shallow rooting and requires nutrient-rich soils and adequate moisture. Extensive irrigation systems, the moisture holding capacity of the soil, and an impressive drainage system ensured that moisture levels were maintained, and water was moved from one terrace to another. The nutrient levels were probably maintained on a regular basis by burning of the terrace surface and the addition of camelid dung (see Kemp et al. 2006 for phosphate values), as well as the rapid rate of weathering of the local volcanic tuff. Maize was a component of the diet of some llama (Lama glama) and alpaca (Vicugna pacos) herds, with animal husbandry being interlinked with cultivation on terracing as part of an integrated agro-pastoral system (Finucane et al. 2006; Cadwallader et al. 2012). However, if maize was being consumed by the camelids, we would expect there to be a greater number of Crosses within the terrace horizons, as llamas and alpacas are normally fed on the leaves and corn husks of the maize plant. It is possible therefore, that the camelids were instead left to graze on the wetland surface and surrounding grasslands, especially as a high proportion of Poaceae phytoliths were found within the terrace deposits, which may have become incorporated into the sequence from the addition of camelid dung. Interestingly, maize phytoliths are also present in the 3bBt1 horizon, representing the pre-terrace ancient land-surface. These could either represent translocation of phytoliths within soil from the 2bABt or cultivation prior to terrace construction during or before the Middle Horizon.
The significance of terrace construction during the Middle Horizon (ad 600–1000), and reconstruction during the late, Late Intermediate Period (ad ~ 1200–1400) (see Branch et al. 2007) are re-evaluated here in light of the new data: (a) age model for the Ayapampa wetland; (b) analysis of the TOC and INF terraces, (c) wealth of palaeoclimatic data for Peru published in about the last 15 years (Rein et al. 2005; Bird et al. 2011; Vuille et al. 2012; Kanner et al. 2013; Apaéstegui et al. 2014; Bustamante et al. 2016; Thompson et al. 2017). The age model from the wetland reveals that sometime between ~ 1,883 to ~ 692 cal bp (ad ~ 67 − 1258; 273 − 242 cm) a thick unit of sandy clay was deposited prior to the renewal of peat formation at ~ 692 cal bp. This period of landscape erosion occurred sometime during the later Early Intermediate Period, Middle Horizon, and early part of the Late Intermediate Period. Despite the chronological uncertainties, and the lack of a precise chronological relationship between the ‘event’ and the cultural history, it is highly likely that the erosion was caused by human induced landscape disturbance given the evidence for: (a) the presence of maize phytoliths at INF and pollen of Solanaceae and Chenopodiaceae/Amaranthaceae in the wetland prior to the Middle Horizon terrace construction, and succeeding this (b) archaeological evidence for widespread Middle Horizon terrace construction, and phytolith evidence for maize cultivation, that would have necessitated significant modification and temporary destabilisation of the land-surface during initial construction. Palaeoclimatic data also indicate that the early Middle Horizon (ad ~ 600–800) was a period of significantly higher precipitation (Fig. 6), which may have led to soil saturation, overland flow of water and accelerated erosion, as well as providing a plentiful water supply for irrigated agriculture. However, the precise cause (or causes) of the ‘event’, and its duration, remain uncertain.

Selected palaeoclimate proxy data for Peru; Huascaran (all records: Thompson et al. 2013), Quelccaya (all records: Thompson et al. 2013), Huagapo Cave (Kanner et al. 2013), Palestina Cave (Apaéstegui et al. 2014), Laguna Pumacocha (Bird et al. 2011), and the Lima 106KL Marine Core (Rein et al. 2005). The key time periods in question for terrace construction and reconstruction (Middle Horizon and Late Intermediate Period) are illustrated by the shaded areas. Maximum age ranges for onset and termination of the Medieval Climate Anomaly (MCA) and Little Ice Age (LIA) are illustrated by shaded bars on the right-hand side of the diagram. Wetter / drier shifts shown by arrows apply to all palaeoclimate graphs. Data downloaded from the National Oceanic and Atmospheric Administration (NOAA), USA Department of Commerce (https://www.noaa.gov/). Data plotted in C2 1.7.7 and edited in Illustrator 2021
Climate change has been heavily debated as the cause for the demise of the Wari Empire at the end of the Middle Horizon. Palaeoclimatic records indicate declining precipitation levels from ad ~ 800–1000 (later Middle Horizon) and ad ~ 1000–1300 (early Late Intermediate Period) suggesting a sustained period of aridity for ~ 500 years during the Medieval Climate Anomaly (Bird et al. 2011; Vuille et al. 2012; Kanner et al. 2013; Apaéstegui et al. 2014). Whatever the cause of the Wari ‘collapse’ (see below) it is reasonable to conclude that terrace abandonment occurred during this period of late Middle Horizon aridity. These conditions were succeeded by more variable climate from the late, Late Intermediate Period (from ~ 650 cal bp; ad ~ 1300) thought to be due to the increased frequency and magnitude of ENSO, or changes in the influence of the South America Summer Monsoon and Intertropical Convergence Zone at the onset of the Little Ice Age (Bird et al. 2011; Kanner et al. 2013; Apáestegui et al. 2014). The renewal of peat formation at ~ 692 cal bp in the Ayapampa wetland broadly coincides with this climatic transition, as does the reconstruction of the terraces during the late, Late Intermediate Period, which may have been a response to higher precipitation during this time.
The archaeological evidence presented earlier suggests however that the ‘collapse’ of the Wari heartland at the end of the Middle Horizon had little direct effect on communities within the Chicha-Soras Valley with only partial abandonment of settlements (Meddens and Branch 2010). Indeed, the onset of the Late Intermediate Period seemingly witnessed significant population growth, with a further demographic shift in the late, Late Intermediate Period (ad ~ 1200–1476). Given the apparent evidence for continuity of occupation (Middle Horizon to Late Intermediate Period transition), it is perhaps surprising that agricultural terraces were seemingly abandoned. Reduced precipitation during the Medieval Climate Anomaly would certainly have had a sustained impact on an agricultural system dependent upon precipitation for irrigation and may have necessitated adaptation in farming practices to mitigate the economic impact. The restoration of terrace farming during the late, Late Intermediate Period, and the notable cultural differentiation between the earlier and later parts of the Late Intermediate Period, suggest an economic response to social and probably climatic changes, which persisted in the Late Horizon. What is urgently required is data from archaeological sites of early and late, Late Intermediate Period age, especially zoological, botanical, and human remains, to test this hypothesis.
Long-distance transfer of plant taxa in the southern-central Peruvian Andes
The presence of Arecaceae and Marantaceae phytoliths in the irrigated terrace soils is remarkable. Neither of these families have species suitable for cultivation or natural growth at this altitude. The agricultural context is particularly significant as previous finds of these families in archaeological sites in Peru have been restricted to domestic occupation or formal ritual contexts (Perry et al. 2006; Duncan et al. 2009; Branch et al. 2014; Hu 2016). Several phytolith deposition pathways can be suggested for the presence of Arecaceae in TOC and INF: Pathway 1 – local growth and cultivation; Pathway 2 – human transportation of soil containing phytoliths to the terrace; and Pathway 3 – human transportation of parts of the plant either unaltered (e.g. fruit and leaves) or altered (e.g. wooden implements and baskets). There are no macro-botanical records for the use of palm from any of the excavated archaeological sites nearby, but this should not preclude its local cultivation despite the environmental conditions today being less favourable for growth (Pathway 1). Transportation of soil during the Inca cultural period is widely recognised from ethnohistorical sources (e.g. Murúa 1946 [1590–1609]; Ogburn 2004a, b, 2014). However, there are few archaeological records for the movement of soil, especially for the purposes of agriculture; instead, the transportation of soil to ceremonial or ritual sites (e.g. Inca stepped platforms or ‘Ushnu’) is known and has been linked to a possible display of Inca sovereignty and/or worship during the agricultural cycle (Branch et al. 2014). This can be attributed to either Pathway 2 or 3 and, based upon ethnohistorical sources, may highlight the mythical and ritual significance of palm (especially ‘Chonta’) for the Inca, and a possible link with the movement of materials between communities throughout the Inca Empire to sites of ritual significance (Branch et al. 2014). The use of chonta in the tip of agricultural tools has been noted in late prehistoric as well as post conquest contexts, attributed to Pathway 3 (see below). The phytoliths occurring through contamination of the samples has been ruled out due to processing procedures, as outlined in the methodology.
There are several potential genera of Arecaceae that could be represented by the palm signature in INF and TOC, which grow within an Andean setting, albeit at lower elevation than the Chicha-Sora Valley. These include Bactris, Ceroxylon, Euterpe and Hyospathe (Huisman et al. 2018). Euterpe and Ceroxylon were highly unlikely due to having Spheroid echinate phytoliths of a larger size and with more crowded projections than those found within the terrace samples (Huisman et al. 2018). Spheroid echinate phytoliths of Hyospathe are of a similar size and shape to those found within the terraces, although not quite as rounded (see Fig. 1F in Huisman et al. 2018). Hyospathe is also considered a sub-Andean genera that occurs infrequently in Peru (Kahn and Moussa 1994), and therefore an unlikely candidate for the palm deposits encountered in this study.
Although Bactris gasipaes, or chonta de pijuayo (peach palm), mainly produces conical palm phytoliths, this species has significant meaning in Peruvian folklore, and therefore may be a candidate for the occurrence of palm within a high-Andean setting based on our ethnohistorical review. Chonta de pijuayo takes a very prominent position today in the mythology of many people living in lowland tropical rain forests because of its importance in a therapeutic role where traditional healers or curanderos use staffs made of ‘chonta’ (Polia 1988, pp 191–193). Depicted in Middle Horizon iconography in the image of the staff god and in the hands of flying shaman on earlier Paracas textiles, staffs were emblems of office (Arnold and Hastorf 2008, pp 121–123). The staff was important in land and water related rituals, in controlling rain (Arnold and Hastorf 2008, p 124) and in organising labour (Gose 1994). In the community of Sarhua in Ayacucho, it was until recently customary to conceal a piece of ‘chonta’ wrapped up in textile in a pocket of the garment worn by the individual concerned, to protect the person from supernatural and people induced harm (Vivanco Pomacanchari C pers. comm.).
The Peruvian highlands have a high frequency of toponyms that include the word ‘chonta’ in their designations. These include the names of mountaintops, rivers and villages. Such as the examples of the Rio Chonta, a tributary to the Rios Pampas and Apurimac, and the mountain peaks of Cerro Chonta Huilca, Chantachonta and Chontalla, and the archaeological sites of Chontamarca in Izcuchaca; all in Ayacucho. As well as the archaeological sites of Chonta Ranra Punta in the Callejón de Huaylas, and Chontay in the Lurín Valley, and the Laguna de Chonta at 4,850 m a.s.l. in in the upper Chillón Valley. These are but a few among many more similar instances of features in the landscape carrying the name of the palm. Chonta de pijuayo and ‘chunta’, in early Quechua dictionaries, translated as palm (Ricardo 1951 [1561], p 39; González Holguín 1989 [1608], p 122), whilst Bertonio in his 1612 Ayamara dictionary translates the term ‘chunta’ as signifying ‘the tip of the hard pole, which is tied to the weeding hoe’ (1984 [1612], p 92). In a contemporary context, highlights the use of ‘chonta’ for the tip of the chaquitaccla, or foot plough, used in agriculture near Cusco. The ubiquitous presence of this reference to palm outside of its natural environment (lowland tropical rainforest) indicates a long timeline for its importance in these highland areas.
In an archaeological context, evidence for palm in the form of phytoliths of Arecaceae is known from the Inca ushnu platform sites of Condormarca (3,251 m a.s.l.) and Ushnupata (3,550 m a.s.l.) in Ayacucho (Branch et al. 2014). Similarly, at the important highland Inca mountaintop shrine of Huanacauri, the analysis of environmental archaeological samples recorded the presence of palm (Bactris sp.) (Kosiba S pers. comm.). Records of ‘chonta’ from Peru include needles made of ‘chonta’ from Late Horizon burials at Huaca Inquisidor (near Lima) (Cornejo 2002, p 187), rods or staffs of ‘chonta’ wrapped in copper sheeting in Late Horizon burials from Qaqas (~ 3,400 m a.s.l.) near Huanta in Ayacucho (Valdez 2002, p 400), bow and shields made of ‘chonta from tomb complex at Conchopata in Ayacucho (Isbell and Cook 2002, p 284), carbonised chonta lances from Middle Horizon closure deposits at the site of Huari (Ochatoma Paravicino et al. 2015), and Middle Horizon timber rods for spindle whorls made from the trunk of palms from La Real, Arequipa (Coleman and Yépez Álvarez 2012). As the palm phytoliths found within Infiernillo and Tocctoccasa terraces originate from an agricultural context, it is probable that they derived from the timber tips of agricultural tools, either from a foot plough or a hoe. Agriculture was likened to warfare in Andean thinking (Bauer 1996, pp 327–332; van de Guchte 1990, pp 335–338). As noted above, ‘chonta’ was frequently used in weaponry, commonly as a timber for clubs (macana) (McEwan 2006, p 128), bows and the tip of lances and arrows. Its use in a foot plough metaphorically deployed as a weapon by farmers implies ‘confrontation’ with the soil, which was vividly stated by Garcilaso de la Vega as they ‘triumphed over’ and ‘disembowelled the earth’, ‘as valiant soldiers’ (1723 [1609], II, p 133). Garcilaso de la Vega’s account contextualises the use of the (here ‘chonta’ tipped) foot plough as a weapon to assault the field surface on the agricultural ‘battlefield’ in order for its regenerative powers to animate the soil to ensure a good harvest.
The second unexpected finding is that of Spheroid ornate phytoliths characteristic of Marantaceae (arrowroot family) e.g. Maranta arundinacea (arrowroot) or Calathea allouia (leren) (Fig. 4b). The family, which typically grows in lowland rainforests, produces an edible rhizome and is a rich source of starch, and hence carbohydrates. Its nutritional properties therefore are widely recognised despite the difficulties extracting the starch. The history of M. arundinacea domestication is of considerable interest with the earliest records from Panama and Colombia dated to 9,200 − 7,600 cal bp (Piperno 2011). In Colombia, records based upon starch grains and grinding stones indicate use of arrowroot at 1,700 m a.s.l. suggesting either an attempt to cultivate at higher elevation, or its transportation from lowland areas (Piperno and Pearsall 1998). Species of Marantaceae can tolerate less favourable environments, for example, cultivated Calathea allouia is highly tolerant of extreme changes in soil moisture content, both saturated and dry but requires frequent irrigation (Martin and Cabanillas 1976). The presence of Marantaceae in the irrigated terrace soils in the Chicha-Soras Valley is perhaps not surprising therefore and may reflect its local cultivation (Pathway 1). Alternatively, the evidence may indicate transportation of the plant (e.g. fibres and rhizomes), or soil and dung containing Marantaceae phytoliths, from lower elevations and their deposition on the terrace surface e.g. green fertiliser (Pathways 2 or 3).
The Marantaceae phytoliths identified in the Chicha-Soras Valley terraces are thought to most likely reflect arrowroot (M. arundinacea), with Chen and Smith (2013) presenting similar Spheroid ornate (Globular regulate) phytoliths from M. arundinacea reference material, consequently we have conducted a review of previous research to support this hypothesis. Maranta arundinacea has been identified in a preceramic context in Peru from starch grains at Buena Vista (Chillón Valley) dating to ~ 4,400 cal bp (Duncan et al. 2009) and from Waynuña in the Cotahuasi Valley (Arequipa) where starch samples were recovered from a floor in a late preceramic house (Perry et al. 2006). The Buena Vista find derives from the interior of a gourd and the use of arrowroot has been attributed here as a foodstuff in a feasting context. Excavations of raised fields near the town of Cerritos in Ecuador have produced likely M. arundinacea phytoliths with late prehistoric to early colonial dates (Stemper 1993, pp 145, 147, 151). Much closer to the Chicha-Soras Valley arrowroot starch grains have come from flaked stone tools of Late Horizon date from the site of Yanawilka (3,050 − 3,090 m a.s.l. and ~ 4.5 km northwest of Vilcashuaman). The nearest source for arrowroot was estimated to be ~ 80.5 km away (Hu 2016, pp 130–131, 135–137, 167–168). The information on usage of arrowroot in a pre-Hispanic context from the Andes is limited. Current references to its role in Peruvian traditional medicine include its use in a tea for those with a delicate or acid stomach, treatment of alcoholism, diarrhoea, dyspepsia, fever, sprains, stress and in the curing of evil character (e.g. Duke 2009, pp 437–439).
Arrowroot, in contrast to palm, is less obvious to explain as an exotic product within a highland agricultural context. It may be that this tropical rhizome finds its origin in elaborate Andean concepts of ‘ritual mixing’. Notions of ‘unity’ and ‘bringing together’ underlie such practices, which use combinations of ritual items brought from different local ecological zones, and distant geographical environments (Gose 1994; Bolin 1998; Ferreira 2014). These traditions are commonly related with notions of agricultural and herding fertility and productivity. Offerings to high altitude mountain deities frequently included the presence of a range of lowland elements in the ritual mesas being prepared on such occasions where such offerings are made to seek protection and fertility from these chthonic sacred beings (Escalante and Valderrama 2014). The importance of obtaining exotic items from distant sources for ritual use and effectively mixing these with a range of items of more local origin was both of great antiquity and widely used. This can, for example, be demonstrated by the use of Spondylus shell from coastal Ecuador in ceremonial contexts across the Andes in powdered, unmodified and worked form (Murra 1980, pp 139–141).
The occurrence of Arecaceae and Marantaceae phytoliths within Middle Horizon agricultural contexts is arguably less surprising than their presence in deposits of Late Intermediate Period date. The latter was a period characterised both locally, as well as across the central and southern Andes, as a period of intensifying conflict and regional warfare, with decreasing social complexity. Reduced or more structured long-distance exchange networks and journeying at times of conflict would have resulted in an increased dependency on local resources and community networks (Arkush 2017). The evidence presented here confirms the existence of exchange networks across distances of at least 130 km for the Late Intermediate Period. Similar long-distance interactions have also been confirmed elsewhere, such as at the Late Intermediate Period defended settlement at Ayawiri, southwest of Lake Titicaca. Here the presence of obsidian, which originated from the Chivay source, confirms long distance exchange with the Colca Valley over 130 km to the east of Ayawiri (Arkush 2017).
Conclusions
The archaeological excavation of two abandoned prehispanic agricultural terraces (Infiernillo and Tocotoccasa) in the Chicha-Soras Valley (Apurimac) in southern-central Peru revealed the presence of palaeosols. The palaeosols represent soil that developed following construction of agricultural terraces during the Middle Horizon. The soil profiles at the current surface developed following reconstruction of the terraces during the Late Intermediate Period. Phytolith analysis revealed an unexpected presence of Arecaceae (palm family) and Marantaceae (arrowroot family) in both terraces, which has been attributed to local cultivation and/or transportation and use of soil, dung, plant material or implements (made of Arecaceae) on the terrace surfaces. Pollen analysis of a nearby wetland (Ayapampa) did not provide evidence for Arecaceae or Marantaceae. Both phytolith and pollen analysis of the terraces and wetland (respectively) indicated that Zea mays was cultivated locally during the Middle Horizon and Late Intermediate Period, although phytoliths of maize are absent from the wetland record during the Middle Horizon. The presence of Solanaceae and Chenopodiaceae/Amaranthaceae pollen in the wetland may be indicative of cultivation of further important taxa during the Middle Horizon and Late Intermediate Period, which continued into the Late Horizon together with Zea mays. Further research on the terraces and wetland would benefit from the analysis of starch grains, normal alkanes (n-alkanes), Rock-Eval pyrolysis and ancient sedimentary DNA, which may be able to identify additional taxa missing from the phytolith and pollen assemblages and develop our understanding of the environmental history (e.g. Cortella and Pochettino 1994; Haslam 2004; Mills et al. 2017; Parducci 2019).
References
Aleman JC, Canal-Subitani S, Favier C, Bremond L (2014) Influence of the local environment on lacustrine sedimentary phytolith records. Palaeogeogr Palaeoclim Palaeoecol 414:273–283
Apaestegui J, Cruz FW, Sifeddine A et al (2014) Hydroclimate variability of the northwestern Amazon Basin near the Andean foothills of Peru related to the South American Monsoon System during the last 1600 years. Clim Past 10:1967–1981
Arkush EN (2011) Hillforts of the ancient Andes: Colla Warfare, Society, and Landscape. University Press of Florida, Gainesville
Arkush E (2017) The end of Ayawiri: abandonment at an Andean Hillfort Town of the late intermediate period. J Field Archaeol 42:241–257
Arnold DY, Hastorf CA (2008) Heads of state: icons. Power and politics in the Ancient and Modern Andes. Left Coast Press, Walnut Creek
Ball TB, Davis AL, Evett RR, Ladwig JL, Tromp M, Out WA, Portillo M (2016) Morphometric analysis of phytoliths: recommendations towards standardization from the International Committee for Phytolith Morphometrics. J Archaeol Sci 68:106–111
Bauer BS (1996) Legitimization of the state in Inca myth and ritual. Am Anthropol 98:327–337
Bertonio L (1984) [1612] Vocabulario de la Lengua Aymara. Centro de Estudios de la Realidad Económica y Social, Montevideo
Bird BW, Abbott MB, Rodbell DT, Vuille M (2011) Holocene tropical south american hydroclimate revealed from a decadally resolved lake sediment δ18O record. Earth Planet Sci Lett 310:192–202
Bolin I (1998) Rituals of respect: the secret of survival in the high Peruvian Andes. University of Texas Press, Austin
Branch NP, Kemp RA, Silva B, Meddens FM, Williams A, Kendall A, Vivanco Pomacanchari C (2007) Testing the sustainability and sensitivity to climatic change of terrace agricultural systems in the Peruvian Andes: a pilot study. J Archaeol Sci 34:1–9
Branch N, Frouin M, Kemp R, Marini N, Meddens F, Onuora C, Silva B (2014) The Landscape, Environment and Pedo-Sedimentary Context of Inca stepped platforms (‘Ushnu’), Ayacucho, Peru. In: Meddens F, Willis K, McEwan C, Branch N (eds) Inca sacred space: landscape, site and symbol in the Andes. Archetype Publications, London, pp 99–117
Bronk Ramsey C (2008) Deposition models for chronological records. Quat Sci Rev 27:42–60
Bronk Ramsey C (2009) Bayesian analysis of radiocarbon dates. Radiocarbon 51:337–360
Bronk Ramsey C, Lee S (2013) Recent and planned developments of the program OxCal. Radiocarbon 55:720–730
Bustamante MG, Cruz FW, Vuille M et al (2016) Holocene changes in monsoon precipitation in the Andes of NE Peru based on δ18O speleothem records. Quat Sci Rev 146:274–287
Cadwallader L, Beresford-Jones DG, Whaley OQ, O’Connell TC (2012) The signs of maize? A reconsideration of what δ13C values say about palaeodiet in the Andean region. Hum Ecol 40:487–509
Caramancia A, Quilter J, Huaman L, Villanueva F, Morales CR (2018) Micro-remains, ENSO, and environmental reconstruction of El Paraíso, Peru, a late preceramic site. J Archaeol Sci Rep 17:667–677
Castillo Luján F (2019) El Chimú temprano de Huaca de la Luna. In: Prieto G, Boswell A (eds) Actas de la Primera Mesa Redonda de Trujillo: Nuevas perspectivas en la arqueología de los valles de Virú, moche y Chicama. Universidad Nacional de Trujillo, Peru, pp 232–268
Chen ST, Smith SY (2013) Phytolith variability in Zingiberales: a tool for the reconstruction of past tropical vegetation. Palaeogeogr Palaeoclim Palaeoecol 370:1–12
Chevalier A (2008) Early Holocene human adaptation in the central Andes: what the phytoliths can tell us? In: Korstanje MA, Babot MP (eds) Matices Interdisciplinarios en Estudios Fitolíticos y de Otros Microfósiles (Interdisciplinary Nuances in Phytoliths and other Microfossil Studies). BAR International Series, vol S1870. British Archaeological Reports, Oxford, pp 149–160
Coleman R, Yépez Álvarez WJ (2012) Instrumentos textiles: Ruecas. In: Yépez Álvarez WJ, Jennings J (eds) ¿Wari en Arequipa? Análisis de los contextos funerarios de La Real. Museo Arqueológico José Mariá Morante, Universidad Nacional de San Agustín, Arequipa, pp 128–144
Cook AG (2004) Wari art and society. In: Silverman H (ed) Andean Archaeology. Blackwell Publishing, Oxford, pp 146–166
Cornejo M (2002) Sacerdotes y tejedores en la provincia inka de Pachacamac. Bol Arqueol PUCP 6:171–204
Correa Trigoso DE, Gamarra Carranza NV, Chaman Medina ME (2018) Los silicofitolitos y gránulos de almidón en cálculos dentales provenientes de contextos funerarios en Chayhuac An, complejo arqueológico Chan Chan, Trujillo, Perú. R Museu Arq Etn 31:80–95
Cortella AR, Pochettino ML (1994) Starch grain analysis as a microscopic diagnostic feature in the identification of plant material. Econ Bot 48:171–181
de la Garcilaso I (1723) [1609] Commentarios Reales que Traten del Origen de los Incas (Primera parte). Oficina Real y Nicolas Rodriguez Franco, Madrid
Dickau R, Whitney BS, Iriarte J et al (2013) Differentiation of neotropical ecosystems by modern soil phytolith assemblages and its implications for palaeoenvironmental and archaeological reconstructions. Rev Palaeobot Palynol 193:15–37
Dickau R, Iriarte J, Quine T, Soto D, Mayle F (2016) Reconstructing pre-colombian agricultural practices in the bolivian savannah: Stratigraphic and phytolith evidence from raised fields at Campo España, western Llanos de Moxos. Cadernos do LEPAARQ (UFPEL) 13:223–267
Dillehay TD (2017) Where the land meets the sea: fourteen millennia of human history at Huaca Prieta, Peru. University of Texas Press, Austin
Dillehay TD, Eling HH Jr, Rossen J (2005) Preceramic irrigation canals in the Peruvian Andes. Proc Natl Acad Sci USA 102:17241–17244
Dong H, Wei X, Li R, Vachula RS, Tan S, Zhou L, Gan T (2022) Burned phytoliths absorbing black carbon as a potential proxy for paleofire. Holocene 32:442–450
Duke JA (2009) Duke’s handbook of Medicinal plants of Latin America. CRC Press, Boca Raton
Duncan NA (2003) At the edge of the Puna: Archaeological test excavation and sampling for phytolith signatures of ancient corrals at Antibal, Peru. M.A. Thesis, University of Missouri, Columbia
Duncan NA, Pearsall DM, Benfer RA Jr (2009) Gourd and squash artifacts yield starch grains of feasting foods from preceramic Peru. PNAS 106:13,202 – 13,206
Escalante C, Valderrama R (2014) Sacred mountains and rituals in the Andes. In: Meddens F, Willis K, McEwan C, Branch N (eds) Inca Sacred Space: Landscape, site and symbol in the Andes. Archetype Publications, London, pp 147–156
Ferreira F (2014) Ritual mixing’: an ethnographic approach to combination of fills from different origins in Inca ushnu platforms. In: Meddens F, Willis K, McEwan C, Branch N (eds) Inca Sacred Space: Landscape, site and symbol in the Andes. Archetype Publications, London, pp 119–126
Finucane B, Maita Agurto P, Isbell WH (2006) Human and animal diet at Conchopata, Peru: stable isotope evidence for maize agriculture and animal management practices during the Middle Horizon. J Archaeol Sci 33:1766–1776
Fisher EC, Albert R-M, Botha G et al (2013) Archaeological reconnaissance for Middle Stone Age Sites along the Pondoland Coast, South Africa. PaleoAnthropol 2013:104–137
Galeano G, Sanín MJ, Mejía K, Pintaud J-C, Millán B (2008) Novelties in the genus Ceroxylon (Arecaceae) from Peru, with description of a new species. Rev Peru Biol 15:65–72
González Holguín D (1989) [1608] Vocabulario de la Lengua General de todo el Peru Llamada Lengua Qquichua o del Inca. Universidad Mayor de San Marcos, Lima
Goodman-Elgar M (2008) Evaluating soil resilience in long-term cultivation: a study of pre-columbian terraces from the Paca Valley, Peru. J Archaeol Sci 35:3:072–3086
Gose P (1994) Deathly Waters and Hungry Mountains: Agrarian ritual and class formation in an Andean Town. University of Toronto Press, Toronto
Grimm EC (1991–1993) TILIA and TILIAGRAPH. Illinois State Museum, Springfield
Grobman A, Bonavia D, Dillehay TD, Piperno DR, Iriarte J, Holst I (2012) Preceramic maize from Paredones and Huaca Prieta, Peru. Proc Natl Acad Sci USA 109:1755–1759
Handley J (2022) Socio-economic responses to late holocene climate variability and environmental change in the Peruvian Andes. Unpublished PhD thesis. https://doi.org/10.48683/1926.00112049. University of Reading, Reading
Harris DR, Hillman GC (1989) Foraging and farming: the evolution of Plant Exploitation. Routledge, London
Haslam M (2004) The decomposition of starch grains in soils: implications for archaeological residue analyses. J Archaeol Sci 31:1715–1734
Henderson A, Galeano G, Bernal R (1995) Field guide to the palms of the Americas. Princeton University Press, Princeton
Hodgson JM (1976) Soil Survey Field Handbook: describing and sampling soil profiles. Technical Monograph 5. Soil Survey of England and Wales, Harpenden
Hogg AG, Hua Q, Blackwell PG et al (2013) SHCal13 Southern Hemisphere calibration, 0–50,000 years cal bp. Radiocarbon 55:1889–1903
Hu D (2016) Labor under the Sun and the Son: landscapes of control and resistance at Inka and Spanish Colonial Pomacocha, Ayacucho, Peru, Unpublished PhD Thesis edn. University of California, Berkeley
Huisman SN, Raczka MF, McMichael CNH (2018) Palm phytoliths of mid-elevation andean forests. Front Ecol Evol 6:193
International Committee for Phytolith Taxonomy (ICPT) (2019) International Code for Phytolith nomenclature (ICPN) 2.0. Ann Bot 124:189–199
Iriarte J, Glaser B, Watling J et al (2010) Late Holocene Neotropical agricultural landscapes: phytolith and stable carbon isotope analysis of raised fields from french Guianan coastal savannahs. J Archaeol Sci 37:2:984–2994
Isbell WH, Cook AG (2002) A new perspective on Conchopata and the Andean Middle Horizon. In: Silverman H, Isbell WH (eds) Andean Archaeology II: Art, Landscape, and Society. Springer, New York, pp 249–305
Kahn F, Moussa F (1994) Diversity and conservation status of Peruvian palms. Biodivers Conserv 3:227–241
Kanner LC, Burns SJ, Cheng H, Edwards RL, Vuille M (2013) High-resolution variability of the south american summer monsoon over the last seven millennia: insights from a speleothem record from the central Peruvian Andes. Quat Sci Rev 75:1–10
Keeley HCM, Meddens FM (1993) Prehispanic agricultural terrace systems in the Chicha-Soras Valley, Peru. Bull Inst Archaeol 29:121–138
Kemp R, Branch N, Silva B, Meddens F, Williams A, Kendall A, Vivanco C (2006) Pedosedimentary, cultural and environmental significance of paleosols within pre-hispanic agricultural terraces in the southern Peruvian Andes. Quat Int 158:13–22
Kendall A, Rodríguez A (2009) Desarrollo y perspectivas de los sistemas de andenería de los Andes centrales del Perú. Institut français d’études andines, Cuzco
Kimura H (1985) Andean exchange: a view from Amazonia. In: Masuda S, Shimada I, Morris C (eds) Andean Ecology and civilization: an interdisciplinary perspective on Andean Ecological Complementarity. University of Tokyo Press, Tokyo, pp 491–504
Lane K (2014) Water technology in the Andes. In: Selin H (ed) Encyclopaedia of the history of science, technology, and medicine in non-western cultures. Springer, Dordrecht, pp 1–24
Lavallée D, Julien M, Béarez P, Usselmann P, Fontugne M, Bolaños A (1999) Pescadores-recolectores arcaicos del extremo sur peruano. Excavaciones en la Quebrada de los Burros (Tacna, Perú). Primeros resultados 1995–1997. Bull Inst fr études andines 28:13–52
Lavallée D, Julien M, Béarez P et al (2011) Quebrada de los burros: Los primeros pescadores del litoral pacífico en el extremo sur peruano (Quebrada de los burros: first fishermen of the south Pacific coast in the far south of Peru). Chungará (Arica) 43(especial):333–351
Lombardo U, Iriarte J, Hilbert L, Ruiz-Pérez J, Capriles JM, Veit H (2020) Early holocene crop cultivation and landscape modification in SW Amazonia. Nature 581:190–193
Martin FW, Cabanillas E (1976) Leren (Calathea allouia), a little known tuberous root crop of the Caribbean. Econ Bot 30:249–256
McEwan GF (2006) The Incas: new perspectives. ABC-CLIO, Santa Barbara
Meddens F, Branch N (2010) The Wari State, its use of ancestors, rural hinterland, and agricultural infrastructure. In: Jennings J (ed) Beyond Wari walls: Regional Perspectives on Middle Horizon Peru. University of New Mexico Press, Albuquerque, pp 155–170
Meddens F, Vivanco PC (2018) The late intermediate period ceramic traditions of Ayacucho, Apurimac and Huancavelica: current thoughts on the Chanca and other regional polities. Ñawpa Pacha Journal of the Institute of Andean Studies 38:3–56
Mills K, Schillereff D, Saulnier-Talbot É et al (2017) Deciphering long‐term records of natural variability and human impact as recorded in lake sediments: a palaeolimnological puzzle. WIREs Water 4:e1195
Murra JV (1980) The Economic Organization of the Inka State. JAI Press, Greenwich
Murúa M (1946) [1590–1609] Historia del Origen y Genealogía Real de los Reyes Incas del Perú. Instituto Santo Toribio de Mogrovejo, Madrid
Ochatoma Paravicino J, Cabrera Romero M, Mancilla Rojas C (2015) El Área Sagrada de Wari: Investigaciones Arqueológicas en Vagachayoc Moqo. Universidad Nacional San Cristóbal de Huamanga, Ayacucho
Ogburn DE (2004a) Evidence for long-distance transportation of building stones in the Inka Empire, from Cuzco, Peru to Saraguro, Ecuador. Lat Am Antiq 15:419–439
Ogburn D (2004b) Power in stone: the long-distance movement of building blocks in the Inca Empire. Ethnohistory 51:101–135
Ogburn DE (2014) From Cusco to the Four Quarters and Vice Versa: connecting Sacred Spaces in the Inca Empire through the Movement of Stone, Sand and Soil. In: Meddens F, Willis K, McEwan C, Branch N (eds) Inca sacred space: landscape, site and symbol in the Andes. Archetype Publications, London, pp 91–98
Osgood Brooks S (1998) Prehistoric agricultural terraces in the Rio Japo Basin, Colca Valley, Peru. University of Wisconsin-Madison, Madison. Unpublished PhD Thesis
Parducci L (2019) Quaternary DNA: a Multidisciplinary Research Field. Quaternary 2:37
Parr JF (2006) Effect of fire on phytolith coloration. Geoarchaeology 21:171–185
Perry L, Sandweiss DH, Piperno DR, Rademaker K, Malpass MA, Umire A, de la Vera P (2006) Early maize agriculture and interzonal interaction in southern Peru. Nature 440:76–79
Perry L, Dickau R, Zarrillo S et al (2007) Starch fossils and the domestication and dispersal of chili peppers (Capsicum spp. L.) in the Americas. Science 315:986–988
Piperno DR (2001) Phytoliths. In: Smol JP, Birks HJB, Last WM (eds) Tracking Environmental Change Using Lake Sediments, Vol 3: Terrestrial, Algal, and Siliceous Indicators. Springer, Dordrecht, pp 235–251
Piperno DR (2006) Phytoliths: a Comprehensive Guide for Archaeologists and Paleoecologists. AltaMira Press, Lanham
Piperno DR (2009) Identifying crop plants with phytoliths (and starch grains) in Central and South America: a review and an update of the evidence. Quat Int 193:146–159
Piperno DR (2011) The Origins of Plant Cultivation and Domestication in the New World Tropics: patterns, process, and New Developments. Curr Anthropol 52:453–470
Piperno DR, Pearsall DM (1998) The silica bodies of tropical American grasses: morphology, taxonomy, and implications for grass systematics and fossil phytolith identification. Smithsonian Contributions to Botany 85. Smithsonian Institution Press, Washington, pp 1–40
Polia M (1988) Glosario del Curanderismo Andino en el Departamento de Piura, Perú. Anthropologica Del Departamento De Ciencias Sociales 6:177–238
Rein B, Lückge A, Reinhardt L, Sirocko F, Wolf A, Dullo WC (2005) El Niño variability off Peru during the last 20,000 years. Paleoceanogr Paleoclim 20:PA4003
Ricardo A (1951) [1561] Vocabulario y phrasis en la lengua general de los indios del Perú, Llamada quichua y en la lengua española. El más copioso y elegante que hasta agora se ha impresso. En los Reyes. Por Antonio Ricardo. Año de MDLXXXVI (1586). Universidad Nacional Mayor de San Marcos, Instituto de Historia de la Facultad de Letras, Lima
Salvador F, Monerris J, Rochefort L (2014/2015) Peatlands of the Peruvian Puna ecoregion: types, characteristics and disturbance. Mires and Peat 15:1–17
Sayre MP (2010) Life across the river: agricultural, ritual, and production practices at Chavín de Huántar, Perú. Unpublished PhD thesis, University of California, Berkeley
Schreiber KJ (1992) Wari Imperialism in Middle Horizon Peru. Anthropological Papers 87. Museum of Anthropology, University of Michigan, Ann Arbor
Stemper DM (1993) The persistence of prehispanic chiefdoms on the Río Daule, Coastal Ecuador. University of Pittsburgh Memoirs in Latin American Archaeology 7. University of Pittsburgh, Department of Anthropology, Pittsburgh
Strömberg CAE (2003) The origin and spread of grass-dominated ecosystems during the Tertiary of North American and how it relates to the evolution of hypsodonty in equids. Unpublished PhD thesis, University of California, Berkeley
Stuiver M, Polach HA (1977) Discussion; reporting of 14 C data. Radiocarbon 19:355–363
Suarez AC, George JJ (2011) Handbook to life in the Inca world. Facts on File, New York
Thompson LG, Mosley-Thompson E, Davis ME, Zagorodnov VS, Howat IM, Mikhalenko VN, Lin P-N (2013) Annually resolved ice core records of tropical climate variability over the past ~ 1800 years. Science 340:945–950
Thompson LG, Mosley-Thompson E, Davis ME, Porter SE (2017) Ice Core Records of Climate and Environmental Variability in the Tropical Andes of Peru: past, Present and Future. Revista de Glaciares y Ecosistemas de Montañam 3:25–40
Troels-Smith J (1955) Karakterisering af løse jordater (characterization of unconsolidated sediments). Danmarks Geologiske Undersøgelse 4, Raekke 3. Geological Survey of Denmark, Copenhagen
Tykot RH, van der Merwe NJ, Burger RL (2006) The importance of maize in initial period and early horizon Peru. In: Staller J, Tykot R, Benz B (eds) Histories of maize: multidisciplinary approaches to the prehistory, linguistics, biogeography, domestication, and evolution of maize. Academic Press, London, pp 187–197
Valdez LM (2002) Y la tradición continúa: la alfarería de la época inka en el valle de Ayacucho, Perú. Bol Arqueol PUCP 6:395–410
Van de Guchte MJD (1990) “Carving the world”: Inca monumental sculpture and landscape. Unpublished PhD Thesis, University of Illinois at Urbana-Champaign, Champaign
Vuille M, Burns SJ, Taylor BL et al (2012) A review of the south american monsoon history as recorded in stable isotopic proxies over the past two millennia. Clim Past 8:1,309-1,321
Watling J, Iriarte J (2013) Phytoliths from the coastal savannas of french Guiana. Quat Int 287:162–180
Watling J, Saunaluoma S, Pärssinen M, Schaan D (2015) Subsistence practices among earthwork builders: Phytolith evidence from archaeological sites in the southwest amazonian interfluves. J Archaeol Sci Rep 4:541–551
Acknowledgements
We are grateful to the following people for their support during the fieldwork: the inhabitants of the village of Pampachiri (Apurimac), Barbara Silva, Cirilo Vivanco, Douglas Walsh and the late Ann Kendall (The Cusichaca Trust). The project was funded by a NERC small research grant no. NER/B/S/2001/00256 (Branch, Meddens), and AHRC SWW DTP studentship no. AH/L503939/1 (Handley). The authors would like to thank the two anonymous reviewers for their comments, whose suggestions helped improve the manuscript.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by Josephine Handley, Nicholas Branch and Michael Simmonds. The first draft of the manuscript was written by Josephine Handley, Nicholas Branch and Frank Meddens. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
There are no competing interests to report of either a financial or non-financial nature.
Additional information
Communicated by W.D. Gosling.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Handley, J., Branch, N., Meddens, F.M. et al. Pre-Hispanic terrace agricultural practices and long-distance transfer of plant taxa in the southern-central Peruvian Andes revealed by phytolith and pollen analysis. Veget Hist Archaeobot 33, 375–391 (2024). https://doi.org/10.1007/s00334-023-00946-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-023-00946-w