
Abstract
Archaeobotanical remains contribute crucial evidence for shifts in human economy from foraging to farming, understanding early village life and the strategies employed by people in the past to cope with changing environmental conditions. However, differential preservation of plant proxies often leads to the over or under representation of some plant types. This research analyses phytoliths and faecal spherulites to provide new perspectives on human economy at the Neolithic site of Abu Hureyra, N. Syria (∼ 11100–6000 bc) and plant taphonomy by comparing results with those from previous extensively analysed charred plant macro-fossils. This site is of especial importance as one of the earliest and largest pre-pottery Neolithic B farming settlements in the world, however, it was flooded following the construction of the Tabqa dam in the 1970s. This research therefore presents a case study for some of the methods that can be applied to archival material to continue research in areas of high archaeological significance that are no longer accessible. The presence of dung spherulites suggests a background faecal component in sediments throughout the lifespan of the site, and should therefore be considered as a depositional pathway for some of the charred plant macrofossils and phytoliths. Phytolith analyses show that a diverse range of vegetation types were exploited throughout the lifespan of Abu Hureyra, reflecting the site’s favourable position on the border of several ecotones, which likely contributed to its longevity over several millennia.
Similar content being viewed by others


Introduction
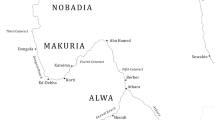
Current research increasingly demonstrates the diversity of the Neolithisation process, pathways and variations between different regions in SW Asia (Fuller et al. 2012; Arranz-Otaegui et al. 2016). There is increasing evidence of highly localised crop practices, community and household scale strategies/adaptations (Bogaard et al. 2017) and the reflection of unique cultural identities of communities through the selection of plant and animal resources (Kabukcu et al. 2021). The Middle Euphrates Valley is an important region for understanding the shift from mobile hunter-gatherers and the development of agricultural settlements (Nesbitt 2002; Willcox et al. 2008; Fuller et al. 2011; Willcox 2012). Tell Abu Hureyra (Fig. 1) is significant for its longevity, with evidence for occupation from ∼ 11100 cal bc as an Epipalaeolithic hunter-gatherer settlement spanning ∼ 1,000 years, and later pre-pottery Neolithic B (PPNB) occupation over several millennia, ∼ 8600 − 6000 cal bc (Hillman 2000a, b, c; Moore et al. 2000, pp 527–529). Abu Hureyra, therefore, provides an important case study for advancing our understanding of the development of early agriculture, sedentism, sustainable environmental management practices, resilience and adaptation to changes in the environment (Roberts et al. 2018).

Location map of Abu Hureyra
Carbonised plant remains provide key evidence for domestication and changes in human economy as the Neolithic developed in SW Asia (van Zeist and Waterbolk-Van-Rooijen 1985; de Moulins 2000; Hillman 2000a, b, c; Hillman et al. 2001; Fairbairn et al. 2002; Willcox 2005; Willcox et al. 2008; Bogaard et al. 2017; Arranz-Otaegui et al. 2018; Weide et al. 2018; Wallace et al. 2019). However, these remains are preserved under a restricted set of conditions and may represent less than 20% of an assemblage compared with desiccated plant remains (van der Veen 2007, p. 977; Dunseth et al. 2019) and sometimes less when compared with micromorphological studies of the diverse plant materials preserved in archaeological deposits (Matthews 2010; Matthews et al. 2020).
Phytoliths are highly durable, as they are inorganic and therefore more resistant to destructive processes than other plant materials (Piperno 2006, p. 5). They are present on many sites globally, particularly in SW Asia, in soils with pH between ∼ 2 and 8 (Weiner 2010, p 175). Phytoliths are microscopic bodies of silica, absorbed in a soluble state by plants through groundwater (Piperno 2006). After the decomposition of organic matter, silica is deposited into soil or sediment, often replicating the cells in which it was deposited in the plants. Following the decomposition of organic matter, the phytoliths are deposited into the soil, providing an indicator of the type of vegetation from which it was derived. Some phytolith morphologies are diagnostic of the parts of the plant in which they formed, for example, the stems and leaves or inflorescences of grasses, and the types and proportions of these can inform on cereal processing activities (Harvey and Fuller 2005; Portillo et al. 2017b) and fuel choices (Gur-Arieh et al. 2013; Portillo et al. 2014, 2017a). Unlike pollen, phytoliths are not usually highly airborne and tend to provide a localised signal of the plants present in a specific deposit and have been used to determine variations in uses of space at archaeological sites (Tsartsidou et al. 2008, 2009; Portillo et al. 2012). Phytoliths are usually identified to the order or family, rather than genus or species level, often providing a lower taxonomic resolution compared with charred macro-fossils. Integration of both proxies, however, provides the potential for a more complete overview of past-plant use (Dunseth et al. 2019). Table 1 highlights some of the key preservation differences between phytoliths and charred macro-fossils as past plant proxies. This study integrates new phytolith data with the previously published charred macro-fossil record from Abu Hureyra (de Moulins 2000; Hillman 2000a, b, c) to provide new perspectives on plant-use and resource management practices during the Neolithic and on plant taphonomy in archaeology more widely.
Knowledge of the depositional and taphonomic processes of plant remains is essential for understanding the wider significance of an assemblage (van der Veen 2007; Matthews 2010). A major challenge in archaeobotany is to disentangle the potential origins of plant material which may contribute to a single context. Animal dung, for example, has been used as fuel since the Epipalaeolithic period in SW Asia (Miller 1984; Miller and Smart 1984; Matthews 2005; Portillo et al. 2014; Spengler 2019; Smith et al. 2019, 2022) to the present day (Miller and Smart 1984; Reddy 1998; Portillo et al. 2017a) and may be a source of seeds and phytoliths as these are preserved in modern and archaeological human and animal coprolites (Shillito et al. 2011; Valamoti 2013; Wallace and Charles 2013; Elliott et al. 2020; Portillo et al. 2020, 2021).
Faecal spherulites from flotation residues suggest the presence of dung at Abu Hureyra during the Epipalaeolithic, and the use of dung fuel from Period 1B (Smith et al. 2022), as suggested by Naomi Miller, who argued that herbivore dung could have contributed to the charred assemblage at Epipalaeolithic Abu Hureyra (Miller 1996). Therefore, increases in small-seeded grasses and legumes, could represent animal diet, rather than a diversification of the human diet in response to environment changes (Hillman et al. 1997) or weed seeds signalling early cultivation (Hillman et al. 2001). Resolving the origins of plant material is key to understanding human subsistence strategies.
During the pre-pottery Neolithic B (PPNB) at Abu Hureyra, charred domesticated cereals and pulses become more prevalent alongside a continued significant presence of small-seeded grasses and legumes (de Moulins 2000; Hillman 2000a, b, c). The identification of dung is important to resolve whether charred macrofossil evidence indicates a continued reliance on a broad-spectrum biodiverse diet of wild, gathered food (Flannery 1969) or whether some of these remains were deposited by animal dung burnt as fuel. A better understanding of whether plant remains represent human food, animal fodder or fuel enables more robust interpretations of the broadening and narrowing of plant-use through different phases of occupation and environmental conditions (Miller 1996).
Furthermore, the presence of dung provides key insights into changing human-animal relationships during the Neolithic, and early stages of animal management, as changes in animal bone morphology indicative of domestication can take up to 1000 years to manifest and may be influenced by environmental and anthropogenic factors (Zeder 2008; Matthews 2010, p. 107; Fuller et al. 2011). Also of major significance and interest is the use of dung as an important secondary product for fuel, manuring or construction (e.g. Bull et al. 2002; Zapata Peña et al. 2003; Matthews 2010; Portillo et al. 2014, 2017a; Gur-Arieh et al. 2019; Dudgeon 2023). Faecal spherulites have been identified in gypsum floor plasters from Abu Hureyra, demonstrating the importance and ubiquity of dung as a resource (Smith et al. 2022; Dudgeon 2023). The use of dung as fuel also provides information on the nature and sustainability of fuel selection and it may also be the preferred fuel choice for specific activities for its long, regular burning properties (Zapata Peña et al. 2003).
However, dung is challenging to identify during excavation as it often appears as amorphous organic material (Shillito et al. 2011) and requires specialised and targeted analytical techniques to detect in archaeological deposits and materials (for overview see Shahack-Gross 2011, p. 206). Dung often disintegrates during flotation and during the phytolith extraction process, resulting in the mixing of plant remains derived from dung with those from other depositional activities (Matthews 2010). One method to assess whether dung is present in archaeological sediments is the identification of faecal spherulites (Matthews 2005; Portillo et al. 2017a; Smith et al. 2019; García-Suárez et al. 2020), microscopic, calcitic particles which form in animals’ guts, particularly ruminants (Brochier et al. 1992; Canti 1997, 1998, 1999).
The key aim of this study is to identify the extent to which dung was used as a resource at Abu Hureyra and was present at the site by analysing faecal spherulites to inform on the likelihood that dung contributed to the fossil plant assemblages and provide new insights into developing human-animal relations. A secondary aim of this study is to analyse phytoliths to provide new perspectives on plant-use at Abu Hureyra.
Study area
Tell Abu Hureyra is located in northern Syria in the Middle Euphrates valley, 35.866°N and 38.400°E, ∼ 130 km east of the modern city of Aleppo (Fig. 1). The Neolithic occupation of Abu Hureyra spans from ∼ 8600 to 6000 cal bc (Fig. 2a) consistent with dates for the mid to late PPNB (Asouti and Fuller 2012, p. 150). The site was built in an area, which is predominately calcareous with a chalk substrate, on a well-drained terrace above the floodplain on the South bank of the river Euphrates, which would have provided a dependable water supply (Moore et al. 2000, p. 28). Abu Hureyra is situated in a key heartland of the development of agriculture, which hosted potentially favourable environmental conditions in the rain-fed agricultural zone and on the banks of the Euphrates. Many cultural developments associated with the Neolithic first occur in the Middle Euphrates and surrounding regions (Akkermans and Schwartz 2003) and through cultural exchange and trade networks, Neolithic Abu Hureyra was linked with contemporary societies in Anatolia, the Southern Levant and Eastern Fertile Crescent (Moore et al. 2000, p. 166). Situated on the border of several ecozones, the inhabitants of Abu Hureyra would have had advantageous access to a broad resource base, which included riverine forest, woodland steppe, stands of wild cereals (wheats, ryes and feather grasses) and park woodland (Moore et al. 2000, pp 43–91).

The site is now flooded and inaccessible following the construction of the Tabqa dam. Professor Andrew Moore and colleagues excavated seven trenches (Fig. 2) during two seasons in 1972 and 1973, revealing densely packed, rectilinear mudbrick buildings across the substantial 11 ha + mound (Moore et al. 1975). However, it is not possible to assess whether all areas of the mound were occupied contemporaneously or represent a series of small settlements over a long period of time. Due to the unique lifespan and significance of the site, and its imminent destruction, an extensive archive of material was recovered (Moore et al. 2000, pp 547–548). Over 100 environmental archaeological bulk samples from the site including soil, occupation residues and charcoal are housed at the University of Reading, providing a unique archive for new environmental analyses.
Economy of Abu Hureyra
The occupation of Abu Hureyra is divided into three periods (Fig. 2a). Abu Hureyra 1 is the Epipalaeolithic settlement, occupied predominately by sedentary hunter-gatherers based on year-round seasons represented in faunal and floral remains. A recent identification of dung spherulites from flotation residues has been used to suggest a small number of animals may have been kept on site and their dung used as an occasional supplementary fuel (Smith et al. 2022).
The Neolithic settlement of Abu Hureyra is characterised by densely packed rectilinear mudbrick buildings, with polished plaster floors, and plaster was also used for vessels and storage containers (Moore et al. 2000, p. 256). The PPNB charred macrofossil assemblage is characterised by high numbers of weed seeds, and the presence of some domesticated type cereals.
In the zooarchaeological assemblage, period 2A is dominated by gazelle, with low numbers of caprines, onager and cattle as well as the sporadic presence of pig and fallow deer. Domesticated cereals including rye, wheat (einkorn and emmer) and barley (two and six rowed) are present, alongside pulses such as lentils, peas and vetches (de Moulins 2000).
The transition between periods 2A and 2B occurred between 7465–7175 cal bc at 95.4% probability (Jacobsson 2017) and is marked by significant settlement growth (∼ 8 ha–∼ 11 ha) and a switch from reliance on wild gazelle to managed caprines in the faunal record (Legge and Rowley-Conwy 2000; Moore et al. 2000, p. 257). There is also a decline in the representation of onager in period 2B, while pig and fallow deer become rare with an increase in cattle exploitation.
In Trench B, there is a slight increase in cereal representation in period 2B, compared with period 2A, accompanied by a decrease in the numbers of samples containing weed seeds. In Period 2B, Trench E, the numbers of weed seeds continue to dominate the assemblage, though cereals are relatively low in number, there is also a gradual reduction in small-seeded legumes (de Moulins 2000).
Materials and methods
Thirty-six samples, selected for phytolith and spherulite analysis were sub sampled from bulk sediments (Table 2). Collected during excavations in the 70s, the bulk samples largely consist of occupation residues, primarily to recover material for radiocarbon dating, and therefore mostly include charred, ashy material, particularly fragments of charcoal. Samples included in this study span the Epipalaeolithic (n = 3), Period 2A (n = 11) and Period 2B (n = 22), with a focus on detecting changes in economy as the PPNB settlement developed. Five of the available samples were included in this study, as corresponding charred macrofossil have been analysed from the same levels, and therefore provide and opportunity to compare the representation of plants through different proxies. Additional contextual information and sample descriptions are provided in ESM 1. Further contextual information including matrices, section drawings and plans are available in Moore et al. (2000, pp 105–131, 189–259).
AH 1 A-C—trench E
Three samples are from the Epipalaeolithic period of occupation at Abu Hureyra (11200–9800 cal bc). E55.31 and E435.15 are fill/occupation residues from pits dating to the earliest phase of occupation, Period 1A (11200–10850 cal bc). Sample E402.14, also representing occupation residues, is dated to the later Epipaleolithic, Period 1C (10850–9800 cal bc). All three samples were brown (10YR 4/3 to 10YR 5/3) silty and slightly ashy sediments.
AH 2A—trench B
Trench B was excavated along the north-south axis of the site (Fig. 2b) and represents a particularly significant area of the excavation because it documents sequential occupation spanning periods 2A, 2B and 2C (Fig. 2a), through 11 phases of building (Moore et al. 2000). The change from reliance on wild gazelle to managed caprine occurred during Trench B, phase 8 (7465–7175 cal bc at 95.4% probability, Jacobsson 2017) and is therefore particularly significant as it demonstrates that the change occurs over time rather than reflecting use of different spaces (Legge and Rowley-Conwy 2000, p. 434).
A limited number of sediment samples were available for analysis from Trench B. B203.99 and B163.71, both representing internal occupation residues dating to Period 2A, phases 5 and 7 respectively, were analysed because the macrofossil assemblages are available (de Moulins 2000)
AH 2A—trench D, phase 4
Trench D was excavated down the western slope of the Abu Hureyra mound (Fig. 2b), initially to ascertain the extent of the settlement although much of the upper layer had eroded away (Moore et al. 2000, pp 209, 218).
Eight samples are analysed in this study from Trench D, all dated to phase 4 (Fig. 2a). Trench D, phase 4 consists predominately of a large open area, between two buildings, which extended beyond the limit of the trench (Fig. 3a). The deposits in the large areas were ashy and burnt, with high quantities of flint and bone, suggesting the intensive use of this area for domestic activities such as cooking (Moore et al. 2000, p. 218). Sample D14.28 is material from a pit, dug into a wall, which contained a Bos primigenius skull. The other samples are all occupation residues from the external area. All of the sediment samples were very dark and ashy, with frequent inclusions of bone and charcoal fragments.

Simplified plans showing a plan of Trench D, phase 4 (adapted from Moore et al. 2000, p. 216, Fig. 8.33). All occupation residues analysed from Trench D, phase 4 (except for D14.28) represent successive layers of occupation in the external activity areas, b plan of Trench E, phase 5 (adapted from Moore et al. 2000, p. 233, Fig. 8.51) and c plan of Trench G, phase 2 (adapted from Moore et al. 2000, p. 247, Fig. 8.65). Approximate sampling location of material analysed in this study shown in bold, where applicable Samples are ordered in stratigraphic sequence as dug (youngest at top)
AH 2B—trench A
One sample was included in this study from Trench A, A207.64 (Fig. 2b). Very little contextual information is currently published from Trenches A or C. This sample was selected as a control, and was sampled from a fragment of mudbrick. There are some limitations to using mudbrick as a control sample, however, in the absence of offsite control samples for the site, the mudbrick provides a baseline for comparing the other samples.
AH 2B—trench E, phases 4–7
During the earliest phase of Neolithic occupation identified in Trench E, phase 4, three rectangular mudbrick houses were identified (Moore et al. 2000, p. 231, Fig. 8.49). The phase 4 buildings set the plan for the construction of buildings throughout period 2B in Trench E (Moore et al. 2000, p. 225). Spaces in between buildings were generally narrow and would have reduced the scope for changing the building plots. The phase 6 buildings were built on the same alignment as the phase 5 buildings (Fig. 3b), however, the house in the centre of the trench in phase 6, had one less room than its phase 5 predecessor, which enlarged the external space between the buildings (Moore et al. 2000, p. 233). During phase 6, the exterior house walls were made much thicker (80 cm long, 30–40 cm wide), although interior walls remained thin (Moore et al. 2000, p. 235). A channel, 30 cm wide and 10 cm deep, filled with ash and charred cultivated chickpea seeds cut between the phase 6 buildings, which excavators hypothesised had been formed by erosion from trampling when animals were herded through the site over a long period of time (Moore et al. 2000, p. 236). Similar to the external areas in Trenches B, D and G, the external areas between houses were filled with lenses of ash and other debris, and were likely the hub of domestic activities, such as cooking (Moore et al. 2000, p. 237).
Sixteen samples are analysed from the Neolithic Period in Trench E, representing phases 5 (n = 10), 6 (n = 5) and 7 (n = 1). Most of the samples represent ashy occupation residues from external areas. E39.33 is from an external hearth base, and E265.76 comes from an external pit filled with ash and occupation debris. Additionally, four of the samples analysed are from internal spaces (Table 2).
AH 2B—trench G, phases 1–3
Trench G was excavated to determine the sequence of occupation in the northeast of the mound (Fig. 2b), the upper parts of which were heavily eroded (Moore et al. 2000, p 241). Five sediment samples from Trench G were analysed for phytoliths and spherulites, representing phases 1–3 from external (n = 2) and internal (n = 2) occupation residues, and an external fire pit (G67.35) (Table 2).
In Phase 1, this area was an open space, with significant deposits of dark occupation soil, patches of burning, with pits and hearths dug into the surface (Moore et al. 2000, p. 242). During phase 2, a mudbrick building was constructed which extended beyond the edge of the trench to the northwest and northeast, of which several rooms were excavated (Fig. 3c). G57.32 represents occupation residues from Room 4 (phase 2), which was an exceptionally narrow room, ∼ 70 cm × 100 cm, with a series of trodden surfaces, renewed as debris built up and possibly used for storage. G62.33 was occupation soil recovered from room 1, where a large number of flint tools were recovered from, including 230 fresh flint blades and lightly retouched blade knives bundled together, but with little flint waste in the deposit (Moore et al. 2000, pp 244, 248). Both G57.32 and G62.33 have corresponding charred macrofossil samples analysed and published which are compared with the phytolith assemblages analysed in this study.
Soil pH
pH was measured for a subset of 16 sediment samples, which were selected to represent different trenches, time periods, deposit and material types to provide an overview of general preservation conditions across the site. Ca. 10 g of air-dried sediment was sieved at 2 mm and weighed into a 50 ml centrifuge tube. 25 ml of ultra-pure water was added using an automatic dispenser. The tube was then capped and placed on an end over shaker working at 20–30 rpm for 15 min. The pH meter was calibrated with pH 7.00 and 9.22 buffers. The pH electrode was placed into the soil suspension, and the pH reading was taken after 30 s. The electrode was cleaned with ultra-pure water between samples to prevent contamination.
Faecal spherulites
The methodology for identifying and quantifying spherulites is based on Canti (1999). Approximately 1 mg of dried sediment was weighed on to a 25 × 75 mm microscope slide, mixed with ∼ 48 µl of clove oil which was distributed evenly over an area of ∼ 22 × 22 mm and cover slipped. The number of spherulites were counted in a known number of fields then related to the initial sediment weight and expressed as number of spherulites per gram of sediment.
Spherulites were identified by size, the presence of a fixed cross of extinction and colour; low order white becomes blue/yellow in opposite quadrants when using the λ plate (Canti 1998) and compared with spherulites derived from modern cow and sheep/goat samples. Spherulites were counted on an optical microscope DMEP at x200 magnification in crossed polarised light (XPL) with further examination at x400 as required. The number of spherulites present in five transects (at x200) were counted.
Phytoliths
Phytoliths were extracted following the rapid extraction method of Katz et al. (2010). Sediments were sieved to remove fractions greater than 0.5 mm and combusted at 500 °C for ∼ 90 min in a muffle furnace to remove organic material. An aliquot of ∼ 40 mg was weighed into a 0.5 ml conical plastic centrifuge tube. 50 µl of 6NHCl was added to dissolve carbonates, followed by 450 µl of Sodium Polytungstate (SPT) (Na6(H2W12WO40)H2O) with a density of 2.4 g/ml to concentrate the phytoliths. The solution was sonicated for 5 min then centrifuged for 5 min at 5,000 RPM. Microscope slides were mounted with 50 µl of the supernatant, which represents 10% of the total number of phytoliths in the initially weighed sample and enabled quantitative comparisons between samples. A minimum of 200 phytoliths with diagnostic morphologies were counted per sample in a known number of fields (between 10 and 50) based on the counting method outlined by Katz et al. 2010. Three or more conjoined cells are counted as multicells and the individual cell morphologies noted to identify the plant type or part it originated from. Each multicell was counted as one phytolith and combined with the single cell count to reach 200 phytoliths. Numbers of phytoliths were related to the initial weight of material to provide an estimated number of phytoliths per gram of sediment. Phytoliths were counted using a Leica DMEP optical microscope at x200 magnification and x400 for further morphological analysis. Digital images were recorded using a Leica DFC420 camera and DMPL optical microscope.
Phytolith morphologies were identified using standard published literature (Twiss et al. 1969; Brown 1984; Piperno 1988; Mulholland and Rapp 1992; Rosen 1992; Fredlund and Tieszen 1994; Strömberg 2004; Neumann et al. 2019), the PhytCore online (Albert et al. 2016) and the University of Reading phytolith reference collection. Nomenclature used within this study follows the most recent International Code for Phytolith Nomenclature, ICPN 2.0, (Neumann et al. 2019) where possible, particularly for geometric morphologies. Modern reference studies (Albert et al. 2003, 2008; Tsartsidou et al. 2007; Portillo et al. 2014) were referred to for the interpretation of phytolith morphologies. Key phytolith morphotypes relevant in this study and their vegetative attributions are summarised in a Table in ESM 3, and photomicrographs showing key morphotypes in ESM 4. Phytoliths which could not be identified because of surface pitting and etching caused by dissolution, were recorded as ‘degraded’ which are expressed as a % of the total phytolith assemblage for each sample.
Comparison of charred macrofossil and phytoliths assemblages
The charred macrofossil assemblages have been analysed and published for six samples which came from the same levels as sediment samples available for analysis in this study; E402, D 59, G57, G62 and B163 (de Moulins 2000; Hillman 2000c). Additionally, macrofossil sample B202 is compared with the sediment sample from B203, as the levels are very close stratigraphically and represent similar context types. As the raw count data for the charred macrofossils from Abu Hureyra was unavailable, the bar lengths of the macrofossil records (de Moulins 2000; Hillman 2000c) were measured to obtain the relative proportions of each taxa per sample following a method similar to that described by Colledge and Conolly (2010, p. 130). The taxa were expressed as a percentage of the total identification per sample. The comparison between the charred plant macrofossils and phytoliths focus on Poaceae and Cyperaceae plant families, as both of these families are visible in both the charred and phytolith assemblages.
Results
Microfossil preservation and concentration
The soil pH in all sediments tested was between 6 and 8 (ESM 1), which is favourable for the preservation of calcitic dung spherulites and silica phytoliths. Dung spherulites were identified in 29 out of 36 samples analysed (Fig. 4a). Where present, concentrations of dung spherulites ranged from an estimated 3,200 to 160,000/g of sediment (Table 2; Fig. 4). Most of the spherulite concentrations were classified as “low” (1–5 spherulites identified and < 20,000 spherulites/g of sediment), although concentrations varied between trenches and time periods (Fig. 4b). Overall, the concentrations of spherulites tended to be higher on average in the later Period 2B (∼ 33,509 spherulites/g of sediment, present in 18 out of 21 samples), compared with Period 2 A (∼ 12,166 spherulites/g of sediment, present in 9 out of 11 samples). However, when Period 2B is separated into Trenches E and G, this trend is less apparent (Fig. 4c), as spherulite concentrations are relatively low and comparable between the samples from Period 2A, Trench D (∼ 7,935 spherulites/g of sediment, present in 5 out of 7 samples) and Period 2B, Trench G (∼ 16,160 spherulites/g of sediment, present in 3 out of 5 samples). Faecal spherulites were present in much higher concentrations in the samples from Period 2B, Trench E (38,931/g of sediment, present in 15 out of 16 samples).

a Bar graph showing estimated number of dung spherulites per gram of sediment, b comparative bar graph showing number of samples by period and trench where spherulites were not identified (absent), low (1–5 spherulites identified = 1–20,000/g sediment), moderate (6–15 spherulites = 21,000–40,000/g sediment) and high (16 + spherulites identified = 41,000 + spherulites/g sediment) and c box and whisker plots comparing individual samples from Trenches D, E and G
Phytoliths were identified in all material analysed in this study, though the estimated number of phytoliths per gram of sediment varied considerably between samples from an estimated 310,000 to 7,000,000 phytoliths/g of sediment (Fig. 5a; Table 2). Phytoliths no longer identifiable due to surface pitting and etching, classified as “degraded”, were also present in all samples and made up between 1 and 33% of the total phytolith assemblage (Table 2).

a Bar graph showing estimated number of phytoliths per gram of sediment and b bar graph showing relative proportions of phytoliths from grasses, sedges, dicots and degraded phytoliths
All assemblages were dominated by phytoliths from monocots, particularly grasses (Poaceae) (Fig. 5b). Most of the grasses which were likely derived from C3 Pooid grasses, based on the prevalence of grass silica short cells (GSSCPs) Rondels, Crenates and Trapeziforms, but most samples also contained GSSCP cells most likely derived from Panicoid grasses (GSSCP Bilobates and Polylobates) (Fig. 6a). Phytoliths diagnostic of sedges (Cyperaceae) were identified in 15 out of the 36 samples analysed (Fig. 5b). Grass phytoliths were derived from stems, leaves and inflorescences, although the proportions of each varied between samples (ESM 2). Plant parts represented by grasses were further explored by calculating the percentage of single and multicell phytoliths which were Elongate dendritics which are diagnostic of grass husks (Fig. 6b). All of the samples analysed included some phytoliths most likely derived from woody or herbaceous plants which included phytoliths from the wood/bark as well as dicot leaves (Figs. 5a and 6c).

a Column graph showing phytoliths most likely derived from chloridoids (GSSCP Saddles), Panicoids (GSSCP Bilobates and Polylobates) and Pooids (GSSCP Crenates, Trapeziforms and Rondels, plus some Bilobates cf. Stipae) and b comparative column graph showing % of Elongate single cells which are Elongate dentritic phytoliths and % of multicell Elongate phytoliths which are Elongate dendritic phytoliths and c stacked column graph showing proportion of phytoliths derived from dicot leaves and dicot wood/bark as a percentage of the total phytolith assemblage
Particularly in SW Asia, multicell phytoliths often provide an opportunity to identify plants, particularly grasses and sedges, to a higher taxonomic resolution (see Table in ESM 2). Although some multicell phytoliths (3 or more cells in anatomical connection) were present in all of the samples analysed in this study and made up between 1 and 20% of the total phytolith assemblage (Table 2), the capacity to provide additional taxonomic resolutions was relatively low. All phytolith multicell count data is available in ESM 3. The results of the spherulite and phytolith analysis are summarised by time period and trench below.
AH 1—trench E
Faecal spherulites were identified in E55.31 and E402.14, however, only a single Spherulite was counted in each sample, equating to ca. 3,200 Spherulites/g of sediment (Table 2; Fig. 4). All three Epipalaeolithic samples had relatively high concentrations of phytoliths, 1.6 to 2.6 million/g of sediment (Table 2; Fig. 5a). Phytoliths which were degraded and exhibited surface pitting and etching, and no longer identifiable, were classified as “degraded”, and made up between 3 and 17% of the total phytolith assemblage in the Epipalaeolithic samples (Table 2). Phytoliths most likely derived from grasses (Poaceae) made up between 72 and 93% of the phytolith assemblages (Fig. 5b). All three samples contained a mixture of different grass types; Pooids, Panicoids and Chloridoids, based on the GSSCP morphologies (Fig. 6a). E55.31 had slightly higher proportions of GSSCP Bilobates and Polylobates usually associated with Panicoid grasses (12%), compared with GSSCP Rondels, Crenates and Trapeziforms, commonly associated with Pooid grasses (9%). E435.15 and E402.14 had slightly higher proportions of GSSCP cf. Pooid grasses (Fig. 6a). GSSCP Saddle phytoliths made up a relatively low proportion of the phytolith assemblage (< 3.5%) in all three samples (Fig. 6a). In all three samples, phytoliths had originated from the stems, leaves and inflorescences (ESM 3). The proportion of Elongate dendritic phytoliths usually formed in the husk has been calculated and shows E55.31 had a relatively high proportion of Elongate dendritic phytoliths, > 50% single cells; >75% multicells, (Figs. 6b and 7).

Summary of the relative percentages of key phytolith multicells identified, grouped by key time periods and trenches
Sedge (“hat shape”) phytoliths were present in E55.31 and E402.14 and made up 1 and 2% of the total phytolith assemblage respectively (Fig. 5b). The relative proportions of single and multicell sedge type phytoliths and Bulliform flabellate phytoliths cf. reeds (Phragmites sp.) (Chen et al. 2020) were combined to provide an indicator of wetland plant resources and made up between ∼ 3 and 11% of the total phytolith assemblage, however no other silica microfossils which are also commonly associated with more moist conditions (diatoms and sponge spicules) were identified in any of the samples (ESM 1).
The phytoliths identified from dicots were derived from both dicot leaves and dicot wood/bark (as defined by the morphotypes in Albert et al. 2003), in relatively equal proportions (Fig. 6c).
AH 2A—trench B
Faecal spherulites were identified in both samples, with a relatively high concentration (48,000/g of sediment in B203.99, 6% of which were darkened (Table 2), indicating burning temperatures between 500 and 800 °C (Canti and Nicosia 2018; Portillo et al. 2020).
Most phytoliths identified were indicative of grasses (Poaceae), which made up 76 and 65% of the total phytolith assemblages (Fig. 5a). Based on GSSCP morphologies, Pooid grasses were most common in both assemblages (∼ 12%), but B203.99 also had a relatively high proportion of cf. Panicoid grasses (9%). No GSSCP Saddles were identified in B163.71 and made up less than 1% of the phytolith percentage in B203.99. The proportion of single and multicell Elongate dendritic phytoliths as a percentage of elongate is much higher in B203.99 (∼ 40–49%), compared with B163.71 (∼ 12%) (Fig. 6b).
Although no phytoliths diagnostic of sedges were identified in either Trench B sample, Bulliform flabellates cf. reeds made up 5 and 12% of the phytolith assemblages in B203.99 and B163.71, providing an indicator of wetland type vegetation, along with a low number of diatoms which were present in B203.99 (ESM 3).
Of the phytoliths likely derived from dicots, both Trench B samples had slightly higher proportions of dicot leaves, compared with phytoliths from the wood/bark (Fig. 6c).
AH 2A—trench D, phase 4
Faecal spherulites were present in seven of the Trench D sample in varying concentrations (Table 2; Fig. 4). Two of the samples, D54.71 and D59.84, contained darkened spherulites indicative of higher burning temperatures (Table 2). The majority of phytoliths identified were indicative of grasses, 61 to 82% of the total phytolith assemblage (Fig. 5b). Based on GSSCP morphology, Pooid grasses were most common, although all sample also contained some GSSCP Bilobates and Polylobates indicative of a Panicoid grass origin, 0.4 to 8.5%, (Fig. 6a). GSSCP Saddles were present in five of the samples, and made up less than 3% of the phytolith assemblage (Fig. 6a). The proportions of Elongate dendritic phytoliths varied considerable between samples, ∼ 15–50% of single cells and 4 to 52% of multicells (Fig. 6b).
Sedge phytoliths were present in four of the Trench D samples and made up between 1 and 5% of the total phytolith assemblage. Combined with Bulliform flabellate phytoliths, all of the samples contained some wetland plant indicators, which ranged from ca. 7 to 17% of the total phytoliths assemblage (ESM 3). Diatoms were present in five of the samples, two of which also contained sponge spicules (ESM 1).
Dicot phytoliths made up between 9 and 36% of the total phytolith assemblage (Fig. 5b), although the proportions of dicot leaves derived from the leaves compared with the wood/bark varied considerably between the samples (Fig. 6c).
AH 2B—trench A
No spherulites were identified in A207.64 (Fig. 4), although some other microfossils were observed in cross polarised light, such as starch grains, these were not quantified. The phytolith concentration was relatively low (680,000/g of sediment), though as it is possible the mudbrick was enhanced with some kind of vegetative temper, the phytoliths should be considered related in some way to anthropogenic input, rather than a true reflection of non-anthropogenic soils in the local environment.
AH 2B—trench E, phases 4–7
Faecal spherulites were identified in all but one of the samples (E268.79) in varying concentrations, from 3,400 to 134,000/g of sediment (Fig. 4). Nine of the samples also contained darkened spherulites indicative of higher temperature burning (Table 2).
The majority of phytoliths in all assemblages were derived from grasses (Poaceae), however, the proportions varied considerably between samples from 44 to 90% (Fig. 5b). Similarly, to the material analysed from other trenches, GSSCP Rondels were most frequently observed indicating the dominance of Pooid grasses in the assemblage, except in sample E344.143 which has a higher proportion of GSSCP cf. Panicoids (∼ 20% compared with 8% GSSCP cf. Pooids) (Fig. 6a). The percentages of both single and multicell Elongate dendritic phytoliths varied considerably between samples (Fig. 6b). Both E344.143 and E268.79 had particularly high proportions of multicell Elongate dendritic phytoliths from grass husks (Fig. 6b).
Phytoliths diagnostic of sedges were present in four of the Trench E Neolithic samples, and made up 1 to 2% of the total phytolith assemblage (Fig. 5b). Combined with Bulliform flabellate phytoliths, wetland plant indicators were present in all samples and varied between ∼ 3 and 14% of the total phytolith assemblage, the lowest proportion represented in the hearth base sample E39.33 (ESM 3). Other microfossil indicators of more moist conditions included diatoms, which were present in six of the samples, four of which also contained sponge spicules. Interestingly, the presence of diatoms and sponge spicules wasn’t related to the presence of sedges or reeds (ESM 3).
Corresponding to the differences in the proportion of grass phytoliths between samples, the proportions of dicot phytoliths also varied, and made up between 8 and 28% of the total phytolith assemblage, with both the lowest and highest proportions from external ashy occupation residues (Fig. 5b). The samples with higher overall proportions of dicots tended to also have higher proportions of dicot wood/bark compared with dicot leaves (Fig. 6c).
AH 2B—trench G, phases 1–3
Faecal spherulites were identified in three out of five of the Trench G samples analysed, in low to moderate concentrations, 20,000 to 37,000/g of sediment, two of which also included darkened spherulites (Table 2; Fig. 4). Significantly, G67.35 from the fire pit contained no faecal spherulites.
The material from Trench G is characterised by relatively low concentrations of phytoliths, 310,000 to 460,000/g of sediment, compared with the other material analysed in this study (Fig. 5a). Similarly to the material from Trenches D and E, grasses made up 61 to 76% of the total phytolith assemblage (Fig. 5b). GSSCPs indicative of Pooid grasses made up between ∼ 2 and 17% of the total phytolith assemblage, and in all samples except for G18.9 and G24.15 were more frequent than GSSCPs indicative of Panicoid grasses (Fig. 6a). No GSSCP Saddles were identified in any of the Trench G samples (ESM 3). The percentage of single cell Elongate dendritic phytoliths varied from ∼ 7 to 31%, and 0 to 33% for multicells.
Sedge “hat shape” phytoliths were present in all of the samples analysed from Trench G and made up between 1 and 7% of the total phytolith assemblage (Fig. 5b, ESM 3). Combined with Bulliform flabellate phytoliths, wetland indicators made up between ∼ 5 and 21% of the total phytolith assemblage (ESM 3).
Phytoliths from dicots made up between 13 and 29% of the total phytolith assemblage, which included slightly higher proportions of phytoliths from the wood/bark compared with dicot leaves (Figs. 5b and 6c).
Comparison with macros
Charred macrofossils have been analysed and published for five of the same contexts as phytoliths have been analysed from in this study; E402, D59, G57, G62 and B163. In addition, the charred macrofossil assemblage from B202 is compared with the phytolith assemblage from B203, as these contexts are closely related stratigraphically and are similar in terms of context and deposit type. The integration of both phytolith and charred macrofossil records from the same assemblages enables this study to investigate how plants are represented in these two different proxies at Abu Hureyra.
Faecal spherulites were observed in all five of the samples with corresponding charred macrofossil samples, although in some cases the concentrations were very low (Fig. 8).

Bar chart comparing relative proportions of grasses and sedges from the phytolith and charred macrofossils records (left) and estimated number of dung spherulites per gram of sediment (right)
The relative proportions of grasses (Poaceae) were consistently higher in the phytolith assemblages compared with the charred macrofossil record for all samples. In fact, samples with high relative proportions of grasses in the phytolith assemblages tended to have relatively lower proportions of grasses in the charred macrofossil assemblages.
Sedges (Cyperacaeae), where present, make up low proportions of the overall plant assemblages in both the charred macrofossil and phytolith records. Where sedges are present in the macro botanical record, they are not identified in the phytolith record and vice versa in this study, with the exception of E402, where sedges are present in both.
The Abu Hureyra charred plant macrofossil assemblages are characterised by exceptionally high relative proportions of small-seeded grasses and legumes, interpreted as weeds of agriculture, and evidence of an increased reliance on domesticates as the Neolithic developed. Small-seeded legumes of the tribe Trifolieae, particularly clovers and medicks, make up over 50% of the charred macrofossil assemblage in all of the above samples except for B163 where the amount is ∼ 20%.
Interpretation and discussion
Dung at Abu Hureyra
The identification of faecal spherulites across the majority (80%) of samples within this study indicates that ruminant dung was both present throughout the occupation of Abu Hureyra, and in places, ubiquitous. A similar conclusion was drawn by Smith et al. (2022) through the identification of faecal spherulites from flotation residue dust, which analysed a different set of samples. Combined therefore, these two studies highlight the need to consider dung as a potential depositional pathway for charred plant macrofossils at Abu Hureyra.
Dung in the Epipalaeolithic at Abu Hureyra
Due to the limited number of sediment samples available for analysis from the Epipalaeolithic period of occupation at Abu Hureyra, this paper, focuses on dung use during the Neolithic periods 2A and 2B. However, given the historical significance of the debate on the use of dung fuel at Epipalaeolithic Abu Hureyra (Miller 1996; Hillman et al. 1997), and the new data suggesting, by Period 1C, people were keeping small numbers of animals on site and using their dung as fuel (Smith et al. 2022), the threes samples analysed from AH 1, are briefly discussed.
Two of the Epipalaeolithic flotation residue samples analysed by Smith et al. (2022) correspond to sediment samples analysed from the same levels in this study, E55.31 and E402.15. The positive identification of faecal spherulites in material from the same contexts from both studies corroborates the results. It is beyond the scope of this paper to quantitatively compare the results for the two methods employed for identifying faecal spherulites, however, a comparison of the two techniques, and integration of the results will be a valuable future study.
Dung in the PPNB at Abu Hureyra
The study by Smith et al. (2022) identified an increase in dung in Neolithic Period 2A compared with the preceding Epipalaeolithic. Similarly, the presence of dung spherulites in 9 out of 11 of the samples analysed from Period 2A in this study, corroborates the findings of Smith et al. (2022). Between Periods 2A and 2B, Smith et al. (2022) observe a decrease in spherulites in flotation, which they attribute to changes in animal management practices whereby as the numbers of domesticated animals increased, they were herded or kept further from the site. In contrast, this study identified a slight increase in spherulite concentration in Period 2B, particularly when comparing external areas from Trench D, Period 2A, compared with Trench E, Period 2B. Although Trench G samples from Period 2B, also have a slightly high concentration of spherulites than Trench D, the difference is negligible compared with the Trench E samples which have much higher concentrations of spherulites (Fig. 4).
A possible reason for the increase in spherulite concentrations identified in this study between periods 2A and 2B could be because of the increase in domesticated animals identified in the archaeozoological record (Legge and Rowley-Conwy 2000). The increase in domestic animals would have increased the availability of a reliable and constant source of dung, whether collected on or offsite. The presence of faecal spherulites in non-pyric features, particularly in external areas in Trenches D and E, representing Periods 2A and 2B, could suggest the continued presence of live animals kept on the site during both periods. Even a small number of live animals kept within the site could have generated a significant amount of waste. Dung may have been burnt to reduce noisome odours, which would have also reduced opportunities for zoonotic diseases to spread. Dung may have been harvested for its favourable burning properties.
A study by the author (Dudgeon 2023) has identified faecal spherulites in gypsum floor plasters from PPNB Periods A, B and C at Abu Hureyra, in samples from Trenches B, D and E. While the study found some variations in the concentrations of spherulites, all of the analysed plaster fragments contained very high concentrations (100,000–1,300,000 spherulites/g of sediment) compared to the sediment samples, indicative of the significant and routine input of dung in plasters throughout the occupation of PPNB Abu Hureyra. Therefore, it seems unlikely that the use of dung fuel decreased in Period 2B based on a decrease in convenience or availability if significant quantities of dung were being used routinely in plaster manufacturing. At Aşıklı Höyük, it has been argued that accumulations of dung between buildings encouraged dung to be recycled and incorporated into construction material (Stiner and Kuhn 2016). In Trench E, between phases 5 and 6, the external space between houses was increased, and a narrow channel is formed, hypothesised to have been created by animals led through the site (Moore et al. 2000). The relatively high concentrations of dung spherulites in external areas from phases 5 and 6, support this hypothesis, and suggest that even if animals were not kept on site, they may have regularly been led through. The presence of faecal spherulites in bulk samples, not specifically targeted as dung deposits, indicates dung was likely accumulating in external activity areas between buildings, and therefore might have been a factor in the use of dung in plaster manufacturing at Abu Hureyra (Smith et al. 2022; Dudgeon 2023).
As argued by Hillman et al. (1997), it is clear wood was routinely and frequently used as a fuel, based on the abundance of wood charcoal. The use of dung as a supplemental fuel has been suggested by the presence of dung spherulites from pyric features by Smith et al. (2022). The identification of dung spherulites in a Period 2B hearth base (E39.33) in this study, also points to the use of dung fuel or the burning of dung for other reasons. The absence of dung spherulites in a fire spot from Trench G, also assigned to period 2B, in this study, may suggest that dung burning practices varied across the site and may have fluctuated over time scales not perceptible in the archaeological record, perhaps even reflecting different seasons where animals were kept closer or further from the site. Variations in floor plaster composition (Dudgeon 2023) tentatively suggest household level selection of specific manufacturing practices. Similarly, the variations in the use of dung fuel in pyric features identified by Smith et al. (2022) and in this study, could also suggest that the selection of dung to supplement fuel could have been a choice specific to different households within the community. At Çatalhöyük, for example, variations in plant-use are observed at a household level (Bogaard et al. 2017), and variations in mudbrick composition have also been attributed to differing recipes which reflect human agency and selection at a household level (Love 2012). It is possible that at Abu Hureyra, as a site of a similar period and scale, there too, were opportunities for households to experiment and express individual preferences through fuel selection.
The presence of ruminant dung in all of the samples compared with the charred macrofossil records in this study, which also contain high proportions of weed seeds could indicate that some of these seeds were deposited by animal dung, which was either burnt as fuel or as a waste management strategy. It is particularly important to consider changes in the relative abundance of possible weed seeds, in light of the confirmation of at least a background faecal component of dung across the site, including some burning of dung, as attested by the presence of spherulites in pyric features (Smith et al. 2022 and this study). Changes in proportions of potential weed-seeds, which have been interpreted as an intensification of the use of domestic crops, could in fact represent fluctuations in animal populations moved on/off or around the site if some of the seeds were deposited by animal dung.
Environment and plant use at Abu Hureyra
All of the phytolith assemblages were dominated by phytoliths derived from monocots, specifically, grasses (Poaceae), but with some sedges (Cyperaceae) or other indicators of more moist environments in all samples, as well as phytoliths derived from the wood/bark and leaves of dicots. The variety of morphologies present in each sample attest the heterogeneous nature of the sediments analysed, which, collected as bulk samples likely included plant input from a number of depositional events. Similarly to at Abu Hureyra, ashy phytolith assemblages identified at Sheikh-e Abad and Jani in the Central Zagros also exhibit highly variable compositions of phytolith morphotypes within each sample, though all ashy samples are fairly similar to one another (Shillito and Elliott 2013, p. 197, Fig. 16.9).
The charred macrofossil assemblages were dominated by dicot plants while monocots dominated the phytolith assemblages (Fig. 8). One of the factors contributing to this contrast is that monocotyledon plants produce up to 20 times more phytoliths than dicots, which are therefore underrepresented in the phytolith record (Albert et al. 2006; Tsartsidou et al. 2007). Another significant factor is that archaeological charred plant remains are dominated by fuel and burnt storage contents (Hillman 1981) while phytoliths represent a broader set of plant uses (Table 1). In addition, dicot wood and bark, which may be used for fuel, can be contaminated by up to 40 or 50% by airborne particles which land on the bark (Albert et al. 2003; Tsartsidou et al. 2007). While both the charred macrofossil and phytolith assemblages were sampled from the same contexts, the methods of recovery and sampling strategy were different, and therefore represent different plant uses and depositional pathways. The flot fractions at Abu Hureyra were recovered in a 1 mm mesh and heavy residues in a 3 mm mesh (Moore et al. 1975, p. 55), though modern standards generally use a 0.25 mm mesh to ensure the recovery of all small-seeded grasses, legumes and chaff (Asouti et al. 2018, p. 25). Furthermore, phytolith representation can vary significantly within a single context (Zurro et al. 2009), which is a consideration in this study, as phytoliths were extracted from bulk samples, and are therefore unlikely to be representative of the whole context. The parts of the plants represented are also different in the charred macrofossil assemblage compared with the phytolith assemblage, which may represent different uses of different plant types. For example, although Pooid cereals tend to produce high numbers of phytoliths; experimental studies have shown phytoliths to be absent or very low in number in the cereal grains (Tsartsidou et al. 2007, p. 1,268, Fig. 2e). Therefore, the cereal grains themselves, identifed in the charred macrofossil assemblage at Abu Hureyra (de Moulins 2000, p. 400) are not necessarily synomonous with high numbers of phytoliths.
Samples with higher proportions of dicot phytoliths, especially those derived from the wood/bark, tended to be from ashier samples with higher proportions of charcoal. This material likely reflected wood burnt as fuel, consistent with the abundant wood charcoal from Abu Hureyra, which made up ∼ 90% of all identified charred remains (Hillman et al. 1997). Where present it is common for charcoal to make up high proportions of charred assemblages compared with other charred plant remains (e.g. seeds/chaff). The consistent presence of dicot phytoliths (Fig. 5b), despite being lower phytolith producers (Albert et al. 2003, p. 470; Tsartsidou et al. 2007), attests the importance of woody vegetation alongside grasses and wetland resources. This is supported by the charred macro-fossil record and charcoal records (de Moulins 2000, pp 399–416; Hillman 2000a, c, pp 341–348; Roitel and Willcox 2000, p. 545).
During the Early Holocene from ca. 9700 cal bc (11,650 cal bp), woodland gradually expanded as a result of increased precipitation and rapid warming (Roberts et al. 2018, p. 49). By period 2A, ∼ 8600 cal bc (10,550 cal bp), regional vegetation reconstructions and charred plants in occupation deposits suggest woodland resources were abundant (de Moulins 2000). Dicots make up an average of 11.8% of the phytolith assemblage during this period, a very slight increase from the proportions of dicots represented in the AH1 samples (Fig. 5b). This could be a reflection of the increasingly wetter conditions in the region, demonstrated by the decrease in δ18O isotopes from Lake Zeribar (Stevens et al. 2001) and Lake Van (Wick et al. 2003; Kwiecien et al. 2014). However, more likely, this reflects the compositions of the bulk samples, which contained more fragments of charcoal. A climate anomaly resulting in cooler, drier conditions across much of the Middle East occurred at 7250 cal bc (∼ 9,200 cal bp) (Fleitmann et al. 2008), although its impact was varied (Flohr et al. 2016). There were no significant changes in the types of vegetation identified in the phytolith record in this study between Periods 2A and 2B which represent occupation prior to and following the cooler, drier conditions which occurred at about 7250 cal bc. However, it could have driven the intensification of already practiced cereal and animal agriculture, resulting in the more widespread agricultural practices in Abu Hureyra 2B compared with period 2A.
Based on GSSCP morphologies, grasses from the Pooideae sub-family, associated with temperate climates (Twiss 1992, pp 115–116) were the most ubiquitous. This is consistent with the charred macro botanical record at Abu Hureyra where Pooideae grasses, including cereals; wheat, barley and rye, have been identified (de Moulins 2000, pp 399–416; Hillman 2000a, c, pp 341–348).
GSSCP Bilobates are most commonly formed in Panicoid grasses (Twiss et al. 1969), however, do also form in other grass sub-families, and in some cases can be distinguished based on their three-dimensional shape and morphology (Neumann et al. 2019). GSSCP Bilobates are occasionally produced in Pooid grasses, particularly in Stipa sp., however, tend to be more trapezoidal in cross section compared with Bilobates from in Panicoids which are more flat and symmetrical (Fredlund and Tieszen 1994; Strömberg 2004, p. 258, Fig. 4h). Bilobates formed in the Aristidoideae and Arundinoideae families on the other hand, tend to have longer, more slender shafts, with convex or “Saddle” like lobes in contrast to the larger, straight or semi-rounded lobes and short, wider shafts typical of Bilobates which form in Panicoid grasses. The Bilobates in this study were therefore typically classed as representing Panicoid grasses, except where the morphology, as described above, more closely represented Pooid types.
Although stands of wild grasses of the millet sub family exist in the Middle Euphrates region today, including torpedo grass (Panicum repens) (Moore et al. 2000, p. 72), Panicoid grasses were much rarer during the occupation of Abu Hureyra. In the charred macrofossil record from Abu Hureyra, Panicoid grasses were represented by low proportions of Bristle grass (Setaria sp.). In Trench B, bristle grass was recorded in B202, which corresponds to a relatively high proportion of Panicoid-type phytoliths inn B203 (∼ 9% of the total phytolith assemblage), compared to B163, where bristle grass was absent from the macrofossil record and Panicoid-type phytoliths made up less than 1% of the total phytolith assemblage (Fig. 6a). In D59, bristle grass was present in the charred macrofossil assemblage and Panicoid-type phytoliths made up ∼ 6% of the charred macrofossil assemblage. In contrast, in Trench G, no bristle grass was reported, but Panicoid phytoliths made up 5–6% in both corresponding samples.
Sedges, as a sub-category of monocots, where present (15 out of 36 samples in this study), made up low proportions of the overall plant assemblages in both the charred macrofossil and phytolith records. Where sedges were present in the macro botanical record, they awere not identified in the phytolith record and vice versa in this study, with the exception of E402, where sedges are present in both. As sedges represent a potentially important resource for a variety of uses including building, bedding, basketry and fuel (Rosen 2005; Ryan 2011; Ramsey et al. 2017, 2018), this contrast highlights a key value of adopting a multi proxy approach. One reason for the identification of sedges in phytolith but not charred macrofossil assemblages is that phytoliths do not require burning for preservation. Sedge type phytoliths, identified in G57, G62 and B163, but absent from the macrofossil assemblage were all sampled from internal occupation residues. The sedges could therefore represent construction material or matting which would not have been burnt and therefore seeds and other macro plant remains would not have been preserved.
The development of agriculture has been traditionally associated with an increased reliance on domestic cereals and legumes (Moore et al. 2000; Weiss et al. 2004; Savard et al. 2006), although a more recent synthesis of archaeobotanical data sets from SW Asia argues there is no evidence for a narrowing of the diet in Neolithic agricultural societies (Wallace et al. 2019). The relative ubiquity of dung during the Neolithic at Abu Hureyra, requires further quantitative assessment alongside the charred macrofossils record to ascertain the full significance of its contribution as a depositional pathway. Preliminarily, the results presented in this study highlight the possibility that reliance on domesticated crops did not necessarily intensify between Periods 2A and 2B, as fluctuations in possible weed seeds could have been deposited by domesticated animals which were herded in the diverse surrounding environs to Abu Hureyra.
Conclusions
Abu Hureyra continues to be an important site for understanding the shift from forging to farming and the development of agricultural societies during the PPNB in SW Asia. The identification of faecal spherulites indicate than dung was present on the site and a potential depositional pathway for at least some of the charred plant macrofossils and phytoliths recovered from the site.
The conclusions from analysis of faecal spherulites from flotation residues in a study by Smith et al. (2022) broadly agree with the outcomes of this study of faecal spherulites from bulk sediment samples. Both studies highlight a faecal presence throughout the occupation of Abu Hureyra, and suggest the use of dung fuel, alongside wood. Minor differences between the spherulite data in both studies could be attributed to the analysis of a different set of samples, which therefore represent spatial differences across the site, context specific variations, or relate to the methodological approaches. A quantitative and empirical comparative study, which integrates the two approaches, is an important avenue for research both at Abu Hureyra and at other sites across SW Asia.
As attested in the charred macrofossil assemblage, the phytoliths also indicate that the inhabitants of Abu Hureyra made use of the rich resource base, including park woodland, steppe grasslands and the valley bottom, which was a likely a factor which contributed to the longevity of the site over several millennia and through periods of climate fluctuations. The presence of ruminant dung in samples with high proportions of small-seeded grasses and legumes, classified as weed seeds, requires further investigation, as some of the possible weed seeds, could be derived from animal dung and therefore represent changes practices in animal management rather than the intensification of domestic crops.
Data Availability
All data is available on request from the author and will be deposited in an online repository in due course.
References
Akkermans PMMG, Schwartz GM (2003) The archaeology of Syria: from complex hunter-gatherers to early urban societies (ca. 16000–300 BC). Cambridge University Press, Cambridge
Albert RM, Bar-Yosef O, Meignen L, Weiner S (2003) Quantitative phytolith study of hearths from the natufian and middle palaeolithic levels of Hayonim cave (Galilee, Israel). J Archaeol Sci 30:461–480. https://doi.org/10.1006/jasc.2002.0854
Albert RM, Bamford MK, Cabanes D (2006) Taphonomy of phytoliths and macroplants in different soils from Olduvai Gorge (Tanzania) and the application to Plio-Pleistocene palaeoanthropological samples. Quat Int 148:78–94. https://doi.org/10.1016/j.quaint.2005.11.026
Albert RM, Shahack-Gross R, Cabanes D et al (2008) Phytolith-rich layers from the late bronze and Iron Ages at Tel Dor (Israel): mode of formation and archaeological significance. J Archaeol Sci 35:57–75. https://doi.org/10.1016/j.jas.2007.02.015
Albert RM, Ruíz JA, Sans A (2016) PhytCore ODB: a new tool to improve efficiency in the management and exchange of information on phytoliths. J Archaeol Sci 68:98–105. https://doi.org/10.1016/j.jas.2015.10.014
Arranz-Otaegui A, Colledge S, Zapata L, Teira-Mayolini LC, Ibáñez J (2016) Regional diversity on the timing for the initial appearance of cereal cultivation and domestication in southwest Asia. Proc Natl Acad Sci USA 113:14001–14006. https://doi.org/10.1073/pnas.1612797113
Arranz-Otaegui A, González Carretero L, Roe J, Richter T (2018) Founder crops” v. wild plants: assessing the plant-based diet of the last hunter-gatherers in southwest Asia. Quat Sci Rev 186:263–283. https://doi.org/10.1016/j.quascirev.2018.02.011
Asouti E, Fuller DQ (2012) From foraging to farming in the southern Levant: the development of epipalaeolithic and pre-pottery neolithic plant management strategies. Veget Hist Archaeobot 21:149–162. https://doi.org/10.1007/s00334-011-0332-0
Asouti E, Ntinou M, Kabukcu C (2018) The impact of environmental change on palaeolithic and mesolithic plant use and the transition to agriculture at Franchthi Cave. Greece PLoS ONE 13:e0207805. https://doi.org/10.1371/journal.pone.0207805
Boardman S, Jones G (1990) Experiments on the effects of charring on cereal plant components. J Archaeol Sci 17:1–11. https://doi.org/10.1016/0305-4403(90)90012-T
Bogaard A, Filipovic D, Fairbairn A et al (2017) Agricultural innovation and resilience in a long-lived early farming community: the 1,500-year sequence at neolithic to early Chalcolithic Çatalhöyük, central Anatolia. Anatol Stud 67:1–28. https://doi.org/10.1017/S0066154617000072
Brochier JE, Villa P, Giacomarra M, Tagliacozzo A (1992) Shepherds and sediments: geo-ethnoarchaeology of pastoral sites. J Anthropol Archaeol 11:47–102. https://doi.org/10.1016/0278-4165(92)90010-9
Brown DA (1984) Prospects and limits of a phytolith key for grasses in the central United States. J Archaeol Sci 11:345–368. https://doi.org/10.1016/0305-4403(84)90016-5
Bull ID, Lockheart MJ, Elhmmali MM, Roberts DJ, Evershed RP (2002) The origin of faeces by means of biomarker detection. Environ Int 27:647–654. https://doi.org/10.1016/S0160-4120(01)00124-6
Cabanes D, Shahack-Gross R (2015) Understanding fossil phytolith preservation: the role of partial dissolution in paleoecology and archaeology. PLoS ONE 10:e0125532. https://doi.org/10.1371/journal.pone.0125532
Cabanes D, Weiner S, Shahack-Gross R (2011) Stability of phytoliths in the archaeological record: a dissolution study of modern and fossil phytoliths. J Archaeol Sci 38:2:480–2490. https://doi.org/10.1016/j.jas.2011.05.020
Canti MG (1997) An investigation of microscopic calcareous spherulites from herbivore dungs. J Archaeol Sci 24:219–231. https://doi.org/10.1006/jasc.1996.0105
Canti MG (1998) The micromorphological identification of faecal spherulites from archaeological and modern materials. J Archaeol Sci 25:435–444. https://doi.org/10.1006/jasc.1997.0210
Canti MG (1999) The production and preservation of faecal spherulites: animals, environment and taphonomy. J Archaeol Sci 26:251–258. https://doi.org/10.1006/jasc.1998.0322
Canti MG, Nicosia C-h (2018) Formation, morphology and interpretation of darkened faecal spherulites. J Archaeol Sci 89:32–45. https://doi.org/10.1016/j.jas.2017.11.004
Chen I, Li K-t, Tsang C-h (2020) Silicified bulliform cells of Poaceae: morphological characteristics that distinguish subfamilies. Bot Stud 61:5. https://doi.org/10.1186/s40529-020-0282-x
Colledge S (1991) Investigations of plant remains preserved in epipalaeolithic sites in the Near East. In: Bar-Yosef O, Valla FR (eds) The natufian culture in the levant. International monographs in prehistory, archaeological series, vol 1. Berghahn Books, New York, pp 391–398
Colledge S, Conolly J (2010) Reassessing the evidence for the cultivation of wild crops during the younger Dryas at Tell Abu Hureyra, Syria. Environ Archaeol 15:124–138. https://doi.org/10.1179/146141010X12640787648504
De Moulins D (2000) Abu Hureyra 2: plant remains from the neolithic. In: Moore AMT, Hillman GC, Legge AJ (eds) Village on the Euphrates: from foraging to farming at Abu Hureyra. Oxford University Press, Oxford, pp 399–416
Dudgeon K (2023) Insights into resource management and technological development through microbotanical and geoarchaeological characterisation of floor plasters from neolithic Abu Hureyra, Syria, 8600–6000 cal BC. Quat Int 655:55–68
Dunseth ZC, Fuks D, Langgut D et al (2019) Archaeobotanical proxies and archaeological interpretation: a comparative study of phytoliths, pollen and seeds in dung pellets and refuse deposits at early islamic Shivta, Negev, Israel. Quat Sci Rev 211:166–185. https://doi.org/10.1016/j.quascirev.2019.03.010
Elliott S, Matthews W, Bull I (2020) Early neolithic animal management and ecology: intergrated analysis of faecal material. In: Matthews R, Matthews W, Rasheed Raheem K, Richardson A (eds) The early neolithic of the Eastern Fertile Crescent. Excavations at Bestansur and Shimshara, Iraqi Kurdistan. Oxbow Books, Oxford, pp 353–395
Fairbairn A, Asouti E, Near J, Martinoli D (2002) Macro-botanical evidence for plant use at Neolithic Çatalhöyük south-central Anatolia, Turkey. Veget Hist Archaeobot 11:41–54. https://doi.org/10.1007/s003340200005
Flannery KV (1969) Origins and ecological effects of early domestication in Iran and the Near East. In: Ubbo PJ, Dimbleby GW (eds) The domestication and exploitation of plants and animals. Duckworth Books, London, pp 73–100
Fleitmann D, Mudelsee M, Burns SJ et al (2008) Evidence for a widespread climatic anomaly at around 9.2 ka before present. Paleoceanography 23:PA1102. https://doi.org/10.1029/2007PA001519
Flohr P, Fleitmann D, Matthews R, Matthews W, Black S (2016) Evidence of resilience to past climate change in Southwest Asia: early farming communities and the 9.2 and 8.2 ka events. Quat Sci Rev 136:23–39. https://doi.org/10.1016/j.quascirev.2015.06.022
Fredlund GG, Tieszen LT (1994) Modern phytolith assemblages from the North American great plains. J Biogeogr 21:321–335. https://doi.org/10.2307/2845533
Fuller DQ, Willcox G, Allaby RG (2011) Cultivation and domestication had multiple origins: arguments against the core area hypothesis for the origins of agriculture in the Near East. World Archaeol 43:628–652. https://doi.org/10.1080/00438243.2011.624747
Fuller DQ, Willcox G, Allaby RG (2012) Early agricultural pathways: moving outside the ‘core area’ hypothesis in Southwest Asia. J Exp Bot 63:617–633. https://doi.org/10.1093/jxb/err307
García-Suárez A, Portillo M, Matthews W (2020) Early animal management strategies during the neolithic of the Konya Plain, Central Anatolia: integrating micromorphological and microfossil evidence. Environ Archaeol 25:208–226. https://doi.org/10.1080/14614103.2018.1497831
Gur-Arieh S, Mintz E, Boaretto E, Shahack-Gross R (2013) An ethnoarchaeological study of cooking installations in rural Uzbekistan: development of a new method for identification of fuel sources. J Archaeol Sci 40:4:331–4347. https://doi.org/10.1016/j.jas.2013.06.001
Gur-Arieh S, Madella M, Lavi N, Friesem DE (2019) Potentials and limitations for the identification of outdoor dung plasters in humid tropical environment: a geo-ethnoarchaeological case study from South India. Archaeol Anthropol Sci 11:2683–2698. https://doi.org/10.1007/s12520-018-0682-y
Harvey EL, Fuller DQ (2005) Investigating crop processing using phytolith analysis: the example of rice and millets. J Archaeol Sci 32:739–752. https://doi.org/10.1016/j.jas.2004.12.010
Hillman GC (1981) Reconstructing crop husbandry practices from charred remains of crops. In: Mercer R (ed) Farming practice in british prehistory. Edinburgh University Press, Edinburgh, pp 123–162
Hillman GC (2000a) Abu Hureyra 1: the Epipalaeolithic. In: Moore AMT, Hillman GC, Legge AJ (eds) Village on the Euphrates: from foraging to farming at Abu Hureyra. Oxford University Press, Oxford, pp 327–399
Hillman GC (2000b) Overview: the plant-based components of subsistence in Abu Hureyra 1 and 2. In: Moore AMT, Hillman GC, Legge AJ (eds) Village on the Euphrates: from foraging to farming at Abu Hureyra. Oxford University Press, Oxford, pp 416–422
Hillman GC (2000c) The plant food economy of Abu Hureyra 1 and 2. In: Moore AMT, Hillman GC, Legge AJ (eds) Village on the Euphrates: from foraging to farming at Abu Hureyra. Oxford University Press, Oxford, pp 327–422
Hillman GC, Legge AJ, Rowley-Conwy PA (1997) On the charred seeds from Epipalaeolithic Abu Hureyra: food or fuel? Curr Anthropol 38:651–655
Hillman G, Hedges R, Moore A, Colledge S, Pettitt P (2001) New evidence of Lateglacial cereal cultivation at Abu Hureyra on the Euphrates. Holocene 11:383–393. https://doi.org/10.1191/095968301678302823
Jacobsson P (2017) Radiocarbon evidence for the pace of the M-/L-PPNB transition in 8th millennium BC southwest Asia. Radiocarbon 59:1677–1689. https://doi.org/10.1017/RDC.2017.110
Kabukcu C, Asouti E, Pöllath N, Peters J, Karul N (2021) Pathways to plant domestication in Southeast Anatolia based on new data from aceramic neolithic Gusir Höyük. Sci Rep 11:2112. https://doi.org/10.1038/s41598-021-81757-9
Katz O, Cabanes D, Weiner S, Maeir AM, Boaretto E, Shahack-Gross R (2010) Rapid phytolith extraction for analysis of phytolith concentrations and assemblages during an excavation: an application at tell es-Safi/Gath, Israel. J Archaeol Sci 37:1557–1563. https://doi.org/10.1016/j.jas.2010.01.016
Kwiecien O, Stockhecke M, Pickarski N et al (2014) Dynamics of the last four glacial terminations recorded in Lake Van, Turkey. Quat Sci Rev 104:42–52. https://doi.org/10.1016/j.quascirev.2014.07.001
Legge AJ, Rowley-Conwy PA (2000) The exploitation of animals. In: Moore AMT, Hillman GC, Legge AJ (eds) Village on the Euphrates: from foraging to farming at Abu Hureyra. Oxford University Press, Oxford, pp 423–471
Love S (2012) The geoarchaeology of mudbricks in architecture: a methodological study from Çatalhöyük. Turk Geoarchaeology 27:140–156. https://doi.org/10.1002/gea.21401
Matthews W (2005) Micromorphological and microstratigraphic traces of uses and concepts of space. In: Hodder I (ed) Inhabiting Çatalhöyük: reports from the 1995–99 seasons. Çatalhöyük Research Project 4. BIAA Monograph 38. McDonald Institute for Archaeological Research, Cambridge, pp 355–398
Matthews W (2010) Geoarchaeology and taphonomy of plant remains and microarchaeological residues in early urban environments in the ancient Near East. Quat Int 214:98–113. https://doi.org/10.1016/j.quaint.2009.10.019
Matthews W, García-Suárez A, Portillo M et al (2020) Integrated micro-analysis of the built environment and resource use: high-resolution microscopy and geochemical, mineralogical, phytolith and biomolecular approaches. In: Matthews R, Matthews W, Rasheed Raheem K, Richardson A (eds) The early neolithic of the Eastern Fertile Crescent. Excavations at Bestansur and Shimshara, Iraqi Kurdistan. Oxbow Books, Oxford, pp 265–286
Miller NF (1984) The use of dung as fuel: an Ethnographic example and an archaeological application. Paléorient 10:71–79. https://doi.org/10.3406/paleo.1984.941
Miller NF (1996) Seed eaters of the ancient Near East: human or herbivore? Curr Anthropol 37:521–528. https://doi.org/10.1086/204514
Miller NF, Smart TL (1984) Intentional burning of dung as fuel: a mechanism for the incorporstion of charred seeds into the archaeological record. J Ethnobiol 4:15–28. https://repository.upenn.edu/penn_museum_papers/50
Moore AMT, Hillman GC, Legge AJ (1975) The excavation of tell Abu Hureyra in Syria: a preliminary report. Proc Prehist Soc 41:50–77. https://doi.org/10.1017/S0079497X00010902
Moore AMT, Hillman GC, Legge AJ (2000) Village on the Euphrates: from foraging to farming at Abu Hureyra. Oxford University Press, Oxford
Mulholland SC, Rapp G Jr (1992) A morphological classification of grass silica-bodies. In: Rapp G, Mulholland SC (eds) Phytolith systematics, emerging issues, vol 1. Springer, New York, pp 65–89
Nesbitt M (2002) When and where did domesticated cereals first occur in southwest Asia? In: Cappers RTJ, Bottema S (eds) The dawn of farming in the near east. Studies in early near eastern production, subsistence, and environment, vol 6. Ex Oriente, Berlin, pp 113–132
Neumann K, Strömberg C, Ball T et al (2019) International Code for phytolith nomenclature (ICPN) 2.0. Ann Bot 124:189–199. https://doi.org/10.1093/aob/mcz064
Piperno DR (1988) Phytolith analysis: an archaeological and geological perspective. Academic Press, San Diego
Piperno DR (2006) Phytoliths: a comprehensive guide for archaeologists and paleoecologists. AltaMira Press, Lanham
Portillo M, Valenzuela S, Albert RM (2012) Domestic patterns in the Numidian site of Althiburos (northern Tunisia): the results from a combined study of animal bones, dung and plant remains. Quat Int 275:84–96. https://doi.org/10.1016/j.quaint.2012.01.024
Portillo M, Kadowaki S, Nishiaki Y, Albert RM (2014) Early neolithic household behavior at tell Seker al-Aheimar (Upper Khabur, Syria): a comparison to ethnoarchaeological study of phytoliths and dung spherulites. J Archaeol Sci 42:107–118. https://doi.org/10.1016/j.jas.2013.10.038
Portillo M, Belarte MC, Ramon J et al (2017a) An ethnoarchaeological study of livestock dung fuels from cooking installations in northern Tunisia. Quat Int 431:131–144. https://doi.org/10.1016/j.quaint.2015.12.040
Portillo M, Llergo Y, Ferrer A, Albert RM (2017b) Tracing microfossil residues of cereal processing in the archaeobotanical record: an experimental approach. Veget Hist Archaeobot 26:59–74. https://doi.org/10.1007/s00334-016-0571-1
Portillo M, García-Suárez A, Matthews W (2020) Livestock faecal indicators for animal management, penning, foddering and dung use in early agricultural built environments in the Konya Plain, Central Anatolia. Archaeol Anthropol Sci 12:40. https://doi.org/10.1007/s12520-019-00988-0
Portillo M, Dudgeon K, Allistone G, Aziz KR, Matthews W (2021) The taphonomy of Plant and Livestock Dung Microfossils: an Ethnoarchaeological and Experimental Approach. Environ Archaeol 26:439–454. https://doi.org/10.1080/14614103.2020.1800344
Ramsey MN, Rosen AM, Nadel D (2017) Centered on the wetlands: integrating new phytolith evidence of plant-use from the 23,000-year-old site of Ohalo II, Israel. Am Antiq 82:702–722. https://doi.org/10.1017/aaq.2017.37
Ramsey MN, Maher LA, Macdonald DA, Nadel D, Rosen AM (2018) Sheltered by reeds and settled on sedges: construction and use of a twenty thousand-year-old hut according to phytolith analysis from Kharaneh IV, Jordan. J Anthropol Archaeol 50:85–97. https://doi.org/10.1016/j.jaa.2018.03.003
Reddy SN (1998) Fueling the hearths in India: the role of dung in paleoethnobotanical interpretation. Paléorient 24:61–70. https://doi.org/10.3406/paleo.1998.4677
Roberts N, Woodbridge J, Bevan A et al (2018) Human responses and non-responses to climatic variations during the last glacial-interglacial transition in the eastern Mediterranean. Quat Sci Rev 184:47–67. https://doi.org/10.1016/j.quascirev.2017.09.011
Roitel V, Willcox G (2000) Analysis of Charcoal from Abu Hureyra 1. In: Moore AMT, Hillman GC, Legge AJ (eds) Village on the Euphrates: from foraging to farming at Abu Hureyra. Oxford University Press, Oxford, pp 544–547
Rosen AM (1992) Preliminary identification of silica skeletons from Near Eastern Archaeological Sites: an Anatomical Approach. In: Rapp G, Mulholland SC (eds) Phytolith Systematics, Emerging Issues. Advances in Archaeological and Museum Science, vol 1. Springer, New York, pp 129–147. https://doi.org/10.1007/978-1-4899-1155-1_7
Rosen AM (2005) Phytolith indicators of plant and land use at Çatalhöyük. In: Hodder I (ed) Inhabiting Çatalhöyük: reports from the 1995–99 seasons. Çatalhöyük research project 4. BIAA monograph 38. McDonald Institute for Archaeological Research, Cambridge, pp 203–212
Ryan P (2011) Plants as material culture in the near Eastern Neolithic: perspectives from the silica skeleton artifactual remains at Çatalhöyük. J Anthropol Archaeol 30:292–305. https://doi.org/10.1016/j.jaa.2011.06.002
Savard M, Nesbitt M, Jones MK (2006) The role of wild grasses in subsistence and sedentism: new evidence from the northern Fertile Crescent. World Archaeol 38:179–196. https://doi.org/10.1080/00438240600689016
Shahack-Gross R (2011) Herbivorous livestock dung: formation, taphonomy, methods for identification, and archaeological significance. J Archaeol Sci 38:205–218. https://doi.org/10.1016/j.jas.2010.09.019
Shillito L, Elliott S (2013) Phytolith indicators of plant resource use at Sheikh-e Abad and Jani. In: Matthews R, Matthews W, Mohammadifar Y (eds) The Earliest Neolithic of Iran: 2008 excavations at Sheik-e Abad and Jani. Oxbow Books, Oxford, pp 185–200
Shillito L-M, Bull ID, Matthews W et al (2011) Biomolecular and micromorphological analysis of suspected faecal deposits at neolithic Çatalhöyük, Turkey. J Archaeol Sci 38:1:869–1877. https://doi.org/10.1016/j.jas.2011.03.031
Smith A, Proctor L, Hart TC, Stein GJ (2019) The burning issue of dung in archaeobotanical samples: a case-study integrating macro-botanical remains, dung spherulites, and phytoliths to assess sample origin and fuel use at tell Zeidan, Syria. Veget Hist Archaeobot 28:229–246. https://doi.org/10.1007/s00334-018-0692-9
Smith A, Oechsner A, Rowley-Conwy P, Moore AMT (2022) Epipalaeolithic animal tending to neolithic herding at Abu Hureyra, Syria (12,800–7,800 cal BP): deciphering dung spherulites. PLoS ONE 17:e0272947. https://doi.org/10.1371/journal.pone.0272947
Spengler RN III (2019) Dung burning in the archaeobotanical record of West Asia: where are we now? Veget Hist Archaeobot 28:215–227. https://doi.org/10.1007/s00334-018-0669-8
Stevens LR, Wright HE Jr, Ito E (2001) Proposed changes in seasonality of climate during the Lateglacial and Holocene at Lake Zeribar, Iran. Holocene 11:747–755. https://doi.org/10.1191/09596830195762
Stiner MC, Kuhn SL (2016) Are we missing the “sweet spot” between optimality theory and niche construction theory in archaeology? J Anthropol Archaeol 44:177–184. https://doi.org/10.1016/j.jaa.2016.07.006
Strömberg CA (2004) Using phytolith assemblages to reconstruct the origin and spread of grass-dominated habitats in the great plains of North America during the late Eocene to early miocene. Palaeogeogr Palaeoclimatol Palaeoecol 207:239–275. https://doi.org/10.1016/j.palaeo.2003.09.028
Tsartsidou G, Lev-Yadun S, Albert R-M et al (2007) The phytolith archaeological record: strengths and weaknesses evaluated based on a quantitative modern reference collection from Greece. J Archaeol Sci 34:1:262–1275. https://doi.org/10.1016/j.jas.2006.10.017
Tsartsidou G, Lev-Yadun S, Efstratiou N, Weiner S (2008) Ethnoarchaeological study of phytolith assemblages from an agro-pastoral village in Northern Greece (Sarakini): development and application of a Phytolith Difference Index. J Archaeol Sci 35:600–613. https://doi.org/10.1016/j.jas.2007.05.008
Tsartsidou G, Lev-Yadun S, Efstratiou N, Weiner S (2009) Use of space in a neolithic village in Greece (Makri): phytolith analysis and comparison of phytolith assemblages from an ethnographic setting in the same area. J Archaeol Sci 36:2:342–2352. https://doi.org/10.1016/j.jas.2009.06.017
Twiss PC (1992) Predicted World distribution of C3 and C4 grass phytoliths. In: Rapp G, Mulholland SC (eds) Phytolith Systematics, Emerging Issues. Advances in Archaeological and Museum Science, vol 1. Springer, New York, pp 113–128. https://doi.org/10.1007/978-1-4899-1155-1_6
Twiss PC, Suess E, Smith RM (1969) Morphological classification of Grass Phytoliths. Soil Sci Soc Am J 33:109–115
Valamoti SM (2013) Towards a distinction between digested and undigested glume bases in the archaeobotanical record from neolithic northern Greece: a preliminary experimental investigation. Environ Archaeol 18:31–42. https://doi.org/10.1179/1461410313Z.00000000021
Van der Veen M (2007) Formation processes of desiccated and carbonized plant remains – the identification of routine practice. J Archaeol Sci 34:968–990. https://doi.org/10.1016/j.jas.2006.09.007
Van Zeist W, Waterbolk-cav Rooijen W (1985) The palaeobotany of tell Bouqras, eastern Syria. Paléorient 11:131–147. https://doi.org/10.3406/paleo.1985.4371
Wallace M, Charles M (2013) What goes in does not always come out: the impact of the ruminant digestive system of sheep on plant material, and its importance for the interpretation of dung-derived archaeobotanical assemblages. Environ Archaeol 18:18–30. https://doi.org/10.1179/1461410313Z.00000000022
Wallace M, Jones G, Charles M et al (2019) Re-analysis of archaeobotanical remains from pre- and early agricultural sites provides no evidence for a narrowing of the wild plant food spectrum during the origins of agriculture in southwest Asia. Veget Hist Archaeobot 28:449–463. https://doi.org/10.1007/s00334-018-0702-y
Weide A, Riehl S, Zeidi M, Conard NJ (2018) A systematic review of wild grass exploitation in relation to emerging cereal cultivation throughout the epipalaeolithic and aceramic neolithic of the Fertile Crescent. PLoS ONE 13:e0189811. https://doi.org/10.1371/journal.pone.0189811
Weiner S (2010) Microarchaeology: beyond the visible archaeological record. Cambridge University Press, Cambridge
Weiss E, Wetterstrom W, Nadel D, Bar-Yosef O (2004) The broad spectrum revisited: evidence from plant remains. PNAS 101:9,551-9,555. https://doi.org/10.1073/pnas.0402362101
Wick L, Lemcke G, Sturm M (2003) Evidence of Lateglacial and Holocene climatic change and human impact in eastern Anatolia: high-resolution pollen, charcoal, isotopic and geochemical records from the laminated sediments of Lake Van, Turkey. Holocene 13:665–675. https://doi.org/10.1191/0959683603hl653rp
Willcox G (2005) The distribution, natural habitats and availability of wild cereals in relation to their domestication in the Near East: multiple events, multiple centres. Veg Hist 14:534–541. https://doi.org/10.1007/s00334-005-0075-x
Willcox G (2012) Searching for the origins of arable weeds in the Near East. Veget Hist Archaeobot 21:163–167. https://doi.org/10.1007/s00334-011-0307-1
Willcox G, Fornite S, Herveux L (2008) Early holocene cultivation before domestication in northern Syria. Veget Hist Archaeobot 17:313–325. https://doi.org/10.1007/s00334-007-0121-y
Zapata Peña L, Peña-Chocarra L, Ibáñez Estévez JJ, González Urquijo JE (2003) Ethnoarchaeology in the moroccan Jebala (Western Rif): wood and dung as fuel. In: Neumann K, Butler A, Kahlheber S (eds) Food, fuel and fields: progress in African archaeobotany. Africa praehistorica, vol 15. Heinrich Barth Institut, Köln, pp 163–175
Zeder MA (2008) Domestication and early agriculture in the Mediterranean Basin: Origins, diffusion, and impact. Proc Natl Acad Sci USA 105:11597–11604. https://doi.org/10.1073/pnas.0801317105
Zurro D, Madella M, Briz I, Vila A (2009) Variability of the phytolith record in fisher-hunter-gatherer sites: an example from the Yamana society (Beagle Channel, Tierra del Fuego, Argentina). Quat Int 193:184–191. https://doi.org/10.1016/j.quaint.2007.11.007
Acknowledgements
The material analysed in this study is housed at the School of Archaeology, Geography and Environmental Sciences and The University of Reading, donated to the university by Andrew Moore, and under the care of Roger Matthews and Wendy Matthews. Special thanks to A. Moore for providing provisional phasing for Trenches A and C. I am very grateful for the comments on an earlier manuscript by W. Matthews, who supervised the MSc and PhD this research was conducted as part of. I extend my thanks to technical staff at the University of Reading who supported laboratory work and to Marta Portillo who provided initial training and support in phytolith analysis. I would also like to thank the two anonymous reviewers who provided detailed and constructive feedback on earlier drafts of this manuscript.
Funding
This research was supported by an Arts and Humanities Research Council, South, West and Wales Doctoral Training Partnership (Grant No. AH/L503939/1 to K Dudgeon) and the University of Reading.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The author declares no competing interests.
Additional information
Communicated by A. Fairbairn.
Publisher’s Note
Springer nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dudgeon, K. New perspectives on plant-use at neolithic Abu Hureyra, Syria: an integrated phytolith and spherulite study. Veget Hist Archaeobot 33, 353–373 (2024). https://doi.org/10.1007/s00334-023-00945-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-023-00945-x