
Abstract
The introduction and adoption of livestock played a pivotal role in shaping subsistence strategies of populations in the southern Iberian Peninsula during the Neolithic. However, there is lack of information regarding animal management strategies, such as grazing areas and changes in foddering strategies, and their correlation with the environmental characteristics, type of site and use of the settlement. The study of feeding strategies of domesticates provides a crucial information about the interaction between the management of the environment, husbandry systems and the exploitation of animal products. In this study, we use the archaeozoological data and the δ13C and δ15N stable isotope composition of the faunal bone collagen to understand herding systems and management strategies during the Neolithic in Phases IV and IIIB at El Toro cave (Antequera, Málaga). Archaeozoological and isotopic results revealed diverse husbandry practices and feeding strategies in El Toro cave during the Neolithic. The variability in δ13C and δ15N values suggests the access of domesticates to different grazing areas and foddering strategies. This study contributes new insights into husbandry practices during the Neolithic and opens new perspectives for analysing animal management in mountain areas.
Similar content being viewed by others


Introduction
The introduction of livestock during the early stages of the Neolithic in the Iberian Peninsula brought about significant changes in the subsistence strategies of populations. Currently, available archaeological data suggest a high degree of variability in animal management practices during the Neolithic in the Iberian Peninsula (Altuna and Mariezkurrena 2009; Aura et al. 2005; Castaños 2004; Martín-Socas et al. 2018; Morales and Riquelme 2004; Navarrete and Saña 2017; Navarrete et al. 2017; 2019; 2023; Pérez-Ripoll 2013, 2016; Saña 2013; Saña et al. 2020; Valente 2016). These management strategies include factors such as herd composition, size and production goals (i.e. meat, wool, milk, traction), often involving some level of human control over animal diet to ensure adaptation to local environmental conditions. In this sense, the study of feeding strategies can provide insight into the relationship between environmental management, husbandry systems and the exploitation of animal products.
Archaeological evidence of Neolithic occupation in the south and southeast of the Iberian Peninsula dates back to the second half of the sixth millennium cal BC, suggesting complex and diversified socio-economic dynamics (Aura et al. 2005; Camalich and Martín-Socas 2013; Martín-Socas et al. 1998, 2018; Morales and Martín 1995; Riquelme 1998; Salvatierra 1994). Different management and exploitation strategies are evidence depending on the region, diversity of biogeographic environments, site type (cave or open-air) and function. However, available archaeozoological data in the south of the Iberian Peninsula point to a successful ecological adaptation of domesticates (Martín-Socas et al. 2018). The dichotomy between wild and domestic species during the Neolithic is evident. During the early Neolithic, domestic animals were predominant in caves and open-air sites (e.g. Los Castillejos and La Molaina, Riquelme 1998; El Toro cave, Watson et al. 2004; Nerja cave, Morales and Martín 1995; El Retamar, Caceres 2002) although high percentages of wild species are documented in some cave sites (e.g. Cueva Chica de Santiago, Pellicer and Acosta 1982; Acosta 1995). Among domesticates recovered from early Neolithic sites, domestic caprines (Ovis aries and Capra hircus) have a higher quantitative representation than other domestic species (Bos taurus and Sus domesticus), with sheep being more abundant than goats. In certain cave sites, such as El Toro cave, it is suggested that domesticates and people cohabited (Égüez et al. 2016; Martín-Socas et al. 2004). From the Late Neolithic, the high predominance of domestic species, with a significant increase in cattle and pig remains is documented (e.g. La Dehesilla, Acosta 1987; Acosta and Pellicer 1990; El Toro cave, Watson et al. 2004; Los Castillejos, Riquelme 1998). However, sites such as Cueva del Nacimiento and Valdecuevas still exhibited a significant presence of wild fauna (Morales and Riquelme 2004).
The success in integrating domesticates into economic practices in the south of the Iberian Peninsula during the early Neolithic is evident. Slaughter pattern results suggest a primary focus on obtaining meat, although secondary product exploitation is proposed from the early Neolithic (Acosta and Pellicer 1990; Riquelme 1998; Watson et al. 2004). Evidence also points to complementarity between agricultural and livestock practices (Buxó 1997; Peña-Chocarro et al. 2013; Rovira 2007; Zapata et al. 2005). However, there is a lack of information regarding animal management strategies during the Neolithic, such as grazing areas or changes in feeding and foddering strategies and their correlation with environmental characteristics of the area, site type and settlement use. For example, in the Antequera region, Neolithic sites primarily documented in caves (Martín-Socas et al. 2004, 2018; Perdiguero 1981; Sanchidrián and García 1987, with one exception being the open-air site Piedras Blancas I: García and Wheatley 2009), were associated with seasonal occupations of highly mobile communities (Martín-Socas et al. 2004, 2018). In this context, studies conducted at El Toro cave (Antequera, Málaga) suggested the synchronous cave use for domestic activities and caprines stabling during the Neolithic (Égüez et al. 2016; Martín-Socas et al. 2004). The sequence of the cave’s stratigraphy, featuring two well-defined Neolithic occupations, allows us to study changes in animal feeding strategies across different periods within the Neolithic.
Stable isotope analysis can be used to study aspects of pastoral management strategies, such as grazing and foddering strategies. In general, there is a consensus that the variable contribution of animal and plant macronutrients to animal diet can potentially reflect husbandry practices in the past (Balasse et al. 2015; Fuller et al. 2012; Halley and Rosvold 2014; Madgwick et al. 2012; Hamilton and Thomas 2012; Hammond and O’Connor 2013; Minagawa et al. 2005; Müldner and Richards 2005; Pechenkina et al. 2005). Previous studies have demonstrated the potential of bulk collagen carbon (13C/12C) and nitrogen (15N/14N) stable isotopes to explore different animal management practices in various contexts, for example, reconstructing strategies used in pig management during the prehistory (Balasse et al. 2015, 2016; Madgwick et al. 2013; Navarrete et al. 2017), characterizing livestock strategies in mountain environments (Villalba-Mouco et al. 2018; Navarrete et al. 2023) or inferring feeding strategies of the herds (Madgwick et al. 2023; Makarewicz 2023; Zavodny et al. 2014). Nevertheless, this approach has not yet been applied to explain husbandry practices in the south Iberian Peninsula during the Neolithic.
In this study, we use the archaeozoological data and the δ13C and δ15N stable isotopes data of the faunal remains to characterize herding systems and management strategies in El Toro cave (Antequera, Málaga) during the Neolithic. These will also contribute to evaluating the availability of herd maintenance areas and the implication of livestock strategies in feeding herds.
Materials
The archaeological site of El Toro cave (Antequera, Málaga)
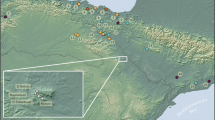
El Toro cave is located in the Sierra del Torcal (Antequera, Málaga) at 1190 m a.s.l. (Fig. 1). Sierra del Torcal is a wide karstic mountain range of calcareous rocks and diaclastic systems that separates Mediterranean Andalusia from the Sub-Baetic System in the southern Iberian Peninsula. The stratigraphic sequence of El Toro cave has been associated with four chrono-cultural phases, of which the lower two correspond to the Neolithic occupations: Phase IV and Phase III (Santana et al. 2019) (Fig. 2).

A Location of El Toro cave (Antequera, Málaga). B General topography of the site of El Toro cave (Antequera, Málaga)

Stratigraphic layers (phases) identified in the site of El Toro cave (Antequera, Málaga)
During Phase IV (Early Neolithic, 6200–5980 BP: 5280–4780 2σ cal BC), the composition and spatial distribution of archaeological remains indicate domestic activities associated with plant and meat processing. Additionally, there is evidence of craft activities such as animal skin treatment and ceramic manufacturing (Martín-Socas et al. 2004; Rodríguez-Rodríguez et al. 1996, 2004). Camalich and Martín-Socas (2013) and Martín-Socas et al. (2004, 2018) interpret Phase IV as a seasonal and periodic occupation. This hypothesis is reinforced by the presence of lithic remains reaching the cave as a finished product (Rodríguez-Rodríguez 2004) and the presence of marine mollusks (Cuenca-Solana et al. 2021). Concerning animal resource management, the predominance of domestic animals over wild is observed. Among domesticates, caprines are predominant, specifically sheep over goats, followed by pigs and cattle (Watson et al. 2004). Micromorphological analysis has identified coprolites from caprines, suggesting that sheep and goats were stabled in the cave (Égüez et al. 2016). The archaeobotanical results record the predominance of wild plants like acorn and wild olives compared to domestic resources like naked wheat (Triticum aestivum), hulled barley (Hordeum distichum), lentils (Lentil sp.), broad beans (Vicia faba) and grass pea (Lathyrus sativus) (Buxó 1997).
Phase III (Late Neolithic, 5320–5170 BP: 4250–3950 2σ cal BC) is subdivided into phases IIIB (base) and IIIA (top) according to the differences in the intensity and spatial distribution of the archaeological remains. At the end of Phase IIIB, structural changes in the cave and the sealing of the main entrance caused by a collapse of the karst system are observed. Micromorphological analysis suggests the synchronous use of the cave as a domestic space and a stable (Égüez et al. 2016). The interior part of the cave would be used for stabling livestock, while the area closer to the entrance was used for domestic and craft activities (Égüez et al. 2016).
Radiocarbon data indicate a more stable and continuous occupation than in Phase IV, which is supported by micromorphological studies (Égüez et al. 2016; Martín-Socas et al. 2004). Domestic cereals such as naked wheat (Triticum aestivum), hulled barley (Hordeum distichum), barley (Hordeum vulgare) and legumes such as lentils (Lentil sp.) and broad beans (Vicia faba) become more representative during this period (Buxó 1997). Regarding livestock, Watson et al. (2004) observed an increase in butchery marks on faunal remains, suggesting a higher level of use and exploitation of animal carcasses. They also noted changes in the slaughter patterns of domestic caprines and could indicate changes in the production and exploitation practices of sheep and goats.
Between both chrono-cultural phases, a sterile deposit was identified and interpreted as a period of site abandonment. Based on dates obtained on short-lived samples, this sterile deposit is estimated to have formed in around 600 years (Martín-Socas et al. 2018).
The selection of faunal remains from Phase IV (early Neolithic) and Phase III (late Neolithic) of El Toro cave
Selection of faunal remains
For this work, the macrofaunal remains of Phase IV and Phase IIIB were reviewed and analysed. A total of 10924 faunal remains were analysed. Considering the main objective of this study is to characterize husbandry strategies, this paper focuses only on the number of identified specimens for Phase IV (NISP = 2132) and Phase IIIB (NISP = 1260) (Table 1).
Selection of faunal samples for isotopic analysis
A total of 82 samples for δ13C and δ15N isotopic analysis in bulk collagen were selected. A range of herbivores (Phase IV n = 28; Phase IIIB n = 21), including domestic (Ovis aries, Capra hircus and Bos taurus) and wild animals (Cervus elaphus, Capra pyrenaica, Capreolus capreolus, Rupicapra rupicapra), omnivores (Phase IV n = 23; Phase IIIB n = 8) (Sus domesticus and Sus scrofa) and carnivores (Phase IV n = 1; Phase IIIB n = 1) (Vulpes vulpes, Canis familiaris) were selected to establish the local δ13C and δ15N isotopic baselines. Samples comprised only adult specimens and included the diaphysis of bones, maxilla and mandibular diastema. Whenever possible, specimens were selected to represent adult individual animals by sampling the same-sided portion of a specific element.
Methods
Archaeozoological analysis
The bones were analysed and identified anatomically and taxonomically in the Museo de Málaga (Spain) using the studies by Boessneck (1980); Fernandez (2001); Gillis et al. (2011); Halstead and Collins (2002); Payne (1985); Prummel and Frisch (1986); Salvagno and Albarella (2017); Zeder and Lapham (2010); and Zeder and Pilaar (2010) to distinguish Rupicapra, Ovis, Capra and Capreolus. The distinction between Sus domesticus and Sus scrofa was carried out on standardized criteria published by von den Driesch (1976) and Payne and Bull (1988). The analysis focused on the representation frequencies of different species and estimate of the age of death of the domesticates (Ovis aries, Capra hircus, Sus domesticus and Bos taurus). The number of identified specimens (NISP) and the minimum number of individuals (MNI) were used to quantify the assemblages. The MNI was calculated by considering the estimated age, anatomical representation and laterality. The relative frequency or presence/absence of taxa was used to estimate taxonomic variability. The age of the animals was estimated according to the stages of tooth wear (Grant 1982; Jones 2006; Lemoine et al. 2014; Payne 1985, 1987), eruption sequences and epiphyseal fusion in postcranial elements (Amorosi 1989; Barone 1976; Silver 1969; Zeder 2006).
Collagen extraction and stable isotope analysis
The extraction of collagen and analysis of stable isotopes were conducted at the Unitat d’Antropologia Biològica (Department of Animal Biology, Plant Biology and Ecology) and Laboratori d’Arqueozoologia (Department of Prehistory) at the Autonomous University of Barcelona (Spain). The bones were cleaned mechanically to remove the surface, and the extraction followed a modified Longin (1971) method (Brown et al. 1988). In short, shards of bones (ca. 200 to 300 mg) were demineralized using 0.6 M HCl at 4°C for several days, then rinsed with ultrapure water (Milli-Q®) and gelatinized with 0.001 M HCl at 80°C for 48 h. Samples were then ultrafiltered (30 kDa, Amicon® Ultra-4 centrifugal filter units; Millipore, MA, USA), frozen and freeze-dried. Collagen samples (0.3 mg) were analysed in duplicate using a Thermo Flash 1112 elemental analyser (EA) coupled to a Thermo Delta V Advantage isotope ratio mass spectrometer (IRMS) with a Conflo III interface at the Institute of Environmental Science and Technology (ICTA) at the Autonomous University of Barcelona (Spain). Isotopic values are expressed as δ. The international laboratory standard IAEA 600 (caffeine) was used as a control. The average analytical error was <0.2‰ (1σ), as determined from the duplicate analyses of δ13C and δ15N. Comparison between δ13C values of wild and domestic herbivores, omnivores and carnivores was performed using one-way ANOVA (α = 0.05) after checking for normal distribution with the Shapiro–Wilk test for normality (α = 0.05). The null hypothesis that the data were normally distributed was rejected for the δ15N values. Thus, the δ15N values were compared using the Kruskal–Wallis test (α = 0.05). All statistical tests were performed in PAST 4.11 (Hammer et al. 2001).
Results
Herd composition
A total of 2132 faunal remains from Phase IV and 1260 faunal remains from Phase IIIB were analysed. The remains of domesticated animals constituted more than 94.4 % (Phase IV, NISP = 2012) and 92.1 % (Phase IIIB, NISP = 1161) of the recovered assemblage, and wild species represented 5.6 % (Phase IV, NISP = 120) and 7.9 % (Phase IIIB, NISP = 99). Among domestic species, in Phase IV and Phase IIIB, caprines were more important quantitatively and in MNI than pig, followed by cattle (Table 1). The results of the comparison of the percentages of each domestic taxon in the Phase IV and Phase IIIIB did not present fluctuations over the period studied.
Mortality profiles were calculated based on the MNI. Pigs and cattle exhibited a similar and homogeneous slaughter pattern across Phase IV and Phase IIIB. Pigs were primarily slaughtered between 14 and 21 months old (Phase IV, 43.5%; Phase IIIB, 57.5%). The number of slaughtered pigs declined after 21 months old (Phase IV, 33.1%; Phase IIIB, 34.4%), and only two individuals documented between 36 and 42 months old in Phase IV (19.5%). Less frequently, neonatal and infantile individual pigs up to 7 months old were documented (Phase IV, 3.9%; Phase IIIB, 8.1%). Regarding cattle, although the total recovered remains were limited, individuals were slaughtered at their meat-optimum (15–30 months old) (Phase IV, 66.7%; Phase IIIB, 66.7%). In both phases, an older individual of more than 48–72 months old was documented (Phase IV, 33.3%; Phase IIIB, 33.3%). The slaughtering patterns of caprines showed slight variations between phases. In both phases, caprine individuals slaughtered between 6 and 24 months old were predominant (Phase IV, Ovis aries 58.9% and Capra hircus 74.4%; Phase IIIB, Ovis aries 62.1% and Capra hircus 78%). However, in Phase IIIB, there was a significant increase in sheep individuals of 2–6 months old (24.4%). The presence of neonates younger than 2 months and infantile of 2–4 months old was also documented in Ovis/Capra during Phase IV (49.8%) and Phase IIIB (40.7%). Distinguishing between sheep and goats in these immature specimens was not feasible due to the high degree of bone fragmentation and the absence of bone development in diagnostic elements.
Faunal stable isotopes δ13C and δ15N values
Collagen preservation
The isotope data and collagen quality indicators are reported in Table 2 and Fig. 3. Out of a total of 82 specimens, collagen was successfully extracted from 69 (84.1%). Collagen yields ranged from 0.3 to 0.4 mg.

Plot of bone collagen δ13C and δ15N values of faunal remains from El Toro cave (Antequera, Málaga). A Phase IV, B Phase IIIB
The C% and N% ranged from 22 to 45% and 11 to 16%, respectively, with C:N ratios ranging from 3.2 to 3.6 and falling within the values proposed by DeNiro (1985) and Van Klinken (1999) for preserved collagen. We also applied a 13% and 4.8% cut-off for C% and N%, respectively, as recommended by Ambrose (1990), Ambrose and Norr (1993).
δ13C and δ15N values
The average δ13C values of herbivores ranged from −19.9 ± 1.2 ‰ (Phase IV; n = 23) to −20 ± 1.1 ‰ (Phase IIIB; n = 18). Although there were no statistically significant differences between phases (p = 0.8345), high intra-range variability was documented: −21.15 to −16.48‰ in Phase IV and −20.88 to −16.48‰ in Phase IIIB. The widest amplitude of variation of δ13C values was recorded in domestic herds (4.85‰) compared to wild animals (1.57‰).
The average δ15N values ranged from +5.2 ± 1 ‰ (Phase IV; n = 23) to +5 ± 1.2 ‰ (Phase IIIB; n = 18). These differences were statistically indistinguishable (p = 0.554). No significant differences were found between the δ13C values of wild and domestic herbivores at Phase IV (p = 0.8408) and Phase IIIB (p = 0.4115). But there were significant statistical differences between δ15N values of wild (average = 4.6 ± 1‰; n = 10) and domestic herbivores (average = 5.6 ± 0.7‰; n = 13) at Phase IV (p < 0.009). Among the domesticated, δ15N values were found to be significantly different between cattle and sheep (p < 0.01) and between cattle and goats (p < 0.03). Cattle were significantly 15N-enriched by 1.3‰ compared to the domestic herbivores (p = 0.007) and 15N-enriched by 2.1‰ compared to the wild herbivores (p = 0.019). In Phase IIIB, there were also significant statistical differences in δ15N values between domestic (average 5.4 ± 1.1 ‰; n = 13) and wild herbivores (average 4.05 ± 0.6 ‰; n = 5) (p = 0.0274). Domesticates were significantly 15N-enriched by 1.35‰ compared to the wild herbivores. Among the domesticated, the only cattle had a δ15N value of 6.89‰. Although there were no statistically significant differences between sheep and goats (p = 0.1366), the δ15N values of sheep ranged from 7.04 ‰ to 3.95 ‰ (n = 9), while the δ15N values of goats ranged from 4.52 to 4.31‰ (n = 3) (Figs. 3 and 4).

Plot of mean δ13C and δ15N values (± 1σ s.d.) of faunal remains from Phase IIIB (blue) and Phase IV (red) of El Toro cave (Antequera, Málaga)
The average δ13C values of pigs ranged from −20.2 ± 0.4 ‰ (Phase IV; n = 19) to −20.8 ± 0.2 ‰ (Phase IIIB; n = 8), and these differences were statistically significant (p < 0.0009). The average δ15N values ranged from +5.7 ± 0.9 ‰ (Phase IV; n = 19) to +5.4 ± 1.1 ‰ (Phase IIIB; n = 8) and were statistically indistinguishable (p = 0.435). In Phase IV, there were no statistically significant differences between pig and wild and domestic herbivores’ δ13C values (wild herbivore, p = 0.202; domestic herbivore, p = 0.377). The pig 15N values were enriched by 0.01‰ compared to the wild herbivores (p = 0.005). Between pig and domestic herbivores, the statistical results of δ15N values were indistinguishable (p = 0.772). In Phase IIIB, δ13C values between pigs and wild herbivores were statistically different (p = 0.025). The δ15N values of pigs were significantly higher by an average of 1.3‰ compared to wild herbivores (p = 0.025). In contrast, there were no statistically significant differences in δ13C and δ15N values between pigs and domestic herbivores (p = 0.890 and p = 0.925, respectively) (Figs. 3 and 4).
Finally, carnivores were only represented by one individual (V. vulpes) from Phase IV. The δ13C value was −18.85‰, and the δ15N value was 7.11‰.
Discussion
Cattle and domestic caprine management strategies
The δ13C values of the specimens analysed in El Toro cave ranged between −21.15 and −16.48‰, reflecting the dominance of C3 carbon in their diets. As expected, the herbivore δ13C values suggest a terrestrial environment dominated by open C3 plants (DeNiro and Epstein 1978; O’Leary 1988). According to a carbon isotope fractionation of ~5‰ between whole plants and consumer’s bone collagen (Ambrose and Norr 1993) and a correction ~ + 1.5‰ due to the fossil fuel effect when applied to pre-industrial ecosystems (Friedli et al. 1986; Hellevang and Aagaard 2015), δ13C values close or higher than −19‰ in herbivorous species could result from the consumption of drought-resistant or dry vegetation, such as shrubs (Filella and Peñuelas 2003). In contrast, plants grown in closed environments produce lower δ13C values (O'Leary 1981; Tieszen 1991). During the early Holocene, the deciduous mesophilic trees such as birch (Betula pendula) and oak (Quercus robur), with more elements of the xerothermophilic maquis and pinewoods coastward, dominated the landscape of the Antequera region (López-Sáez et al. 2011).
The variation of δ13C values δ15N values observed between cattle and domestic caprines serves as an indication that these animals fed on plant resources with diverse isotopic composition. Specifically, cattle exhibit a δ13C range of 4.67‰ and a 15N-enriched by 2.3‰ compared to the wild herbivores. Wild herbivores are generally less restricted in their ecological niche. In Phase IV, cattle have a wider range of δ13C values (−21.1 to −16.48‰) than in Phase IIIB (−16.48‰). However, the amplitude of variation of δ15N values between phases is minimal (0.16‰). The specimens of Phase IV (ID=33) and Phase IIIB (ID= 84) form a distinct isotopic group (Figs. 3 and 4). Two hypotheses can be considered. Firstly, the low representation percentages and MNI of cattle (Phase IV, NISP = 69; MNI = 3; and Phase IIIB, NISP = 48; MNI = 3) compared to the rest of the domestic species suggest that cattle would not be a frequently consumed resource in the cave. The narrow δ13C range of wild herbivores could indicate that these cattle could be tentatively associated with non-local animals, raised elsewhere and brought later to the site. Martín-Socas et al. (1999, 2004) suggest that El Toro cave might have been part of a regional trading network. The authors support this hypothesis by pointing to the recovery of malacofauna, ichthyofauna, raw materials and plant remains that were not sourced from the immediate area of the cave. For example, the recovery of ichthyofauna or malacofauna suggests frequent movements between the mountainous area and the coastal zone, which is approximately 35 km south in a straight line (Martín-Socas et al. 2004). Moreover, shrubs and branches collected over a wide geographical area, around the cave to the basal areas of the mountains, were documented inside the cave (Buxó 1997; Lopez-Saez et al. 2011; Martín-Socas et al. 2004; Rodríguez-Rodríguez et al. 1996). For example, Pistacia sp. and heather (Calluna vulgaris) require decalcified soils or mastic tree that requires areas sheltered from the cold or warmer areas than those existing in the Sierra del Torcal (Martín-Socas et al. 2004).
Secondly, the cattle may have had access to nearby pastures, and the change in δ13C values could be a consequence of the changing environmental conditions. The first signs of anthropization in Sierra de El Torcal are detectable from the second half of the sixth millennium cal. BC and are associated, in many cases, with deforestation caused by fire (Lopez-Saez et al. 2011). In the fifth millennium cal. BC (Phase IIIB), livestock practices significantly transformed the landscape of Sierra de El Torcal, resulting in vegetation degradation and the proliferation of understory and scrub species (Martín-Socas et al. 2018; Rodríguez-Ariza 1996, 2004). Similar transformations have been observed in Cueva de Los Murciélagos (Córdoba) and Polideportivo de Martos (Jaén) (Lizcano 1999; Rodríguez-Rodríguez et al. 1996). Conversely, 15N-enriched in animals may indicate consumption of 15N-enriched plants fertilized with manure (Bogaard et al. 2007; Yousfi et al. 2010; Makarewicz 2014). This 15N-enriched may suggest the consumption of agricultural by-products that were fertilized with manure, a phenomenon documented in various Neolithic contexts across Europe (Bogaard et al. 2013; Fraser et al. 2011). The high values of δ15N could indicate cattle accessing cultivated fields. However, the impact of animal dung on plant δ15N values is highly variable and depends on factors such as manuring frequency and type of fertilizer (Fraser et al. 2011; Szpak 2014). In El Toro cave, cereals such as naked wheat, naked barley and dressed barley and legumes such as lentils, broad beans and pebbles have been documented (Buxó 1997). However, the number of cattle samples from Phase IV (NISP=2) and Phase IIIB (NISP=1) is too small to lead to conclusive interpretations.
Domestic caprines are the most common taxon in El Toro cave during Phase IV (Ovis/Capra NISP = 1195; sheep NISP = 222; goat NISP = 146) and Phase IIIB (Ovis/Capra NISP = 661; sheep NISP = 136; goat NISP = 105), with sheep slightly outnumbering goats. Slaughtering patterns of caprines in Phase IV and Phase IIIB suggest a primary focus on meat production, evidenced by animals being killed between 6 and 24 months old. Still, a significant percentage of animals killed before 6 months old. Such early slaughtering of animals is commonly associated with either milk production for human consumption or the acquisition of high-quality tender meat (Oueslati 2006). Furthermore, in El Toro cave, evidence of dairy consumption has been provided (Tarifa-Mateo et al. 2019).
As already pointed out by Watson et al. (2004), the archaeozoological and microstratigraphic analysis of the site confirmed that caprine penning inside the cave during Phase IV and Phase IIIB (Égüez et al. 2016). The intensity of the cave’s occupation during Phase IV is evidenced by abundant microscopic fragments of charred seeds and ceramics and burning events commonly found in caprine penning contexts (Égüez et al. 2016).
The isotopic analysis of domestic caprines shows a wide range in δ13C values, particularly in sheep (Phase IV = 3.18‰; Phase IIIB = 1.83‰). Goats show differentiated ranges between phases (Phase IV = 0.17‰; Phase IIIB = 2.50‰). A wide range in δ15N values is also documented. The 15N-enriched values of sheep (0.63‰) and goats (0.36‰) were lower than those documented for sheep in Phase IIIB (1.49‰). Conversely, goats maintained a consistently low value in Phase IIIB (0.9‰). These wide ranges in δ13C and δ15N may result from various factors, including different grazing areas, herd movement, foddering strategies or management strategies of stabling areas. If there were seasonal herd movements, they might not be discernible from the analysis of carbon and nitrogen stable isotopes in bone collagen. Three hypotheses are considered. Firstly, the access of flocks to diverse pastures enriched in 15N by manure might explain the variability in δ15N values observed between Phase IV and Phase IIIB. During Phase IV, wild plants constituted 62%, and cultivated plants only constituted 39% of the human diet (Buxó 1997). Phase IIIB represents a systematic occupation of the cave (Martín-Socas et al. 2004), with an intensification of agriculture and an increase in the importance of cultivated species (cereals as wheat, hulled barley, barley and legumes such as lentils and broad beans) versus wild species (acorns) in the cave (Buxó 1997). Differences in isotopic values between sheep and goats could be conditioned by their distinct dietary preferences and needs (Vaiglova et al. 2014), as well as documented grazing habits during early Neolithic at La Draga site (Navarrete et al. 2019). The recorded values for El Toro cave are lower than those documented in the European context (Bogaard et al. 2013). However, it aligns with the values recorded for the northeast of the Iberian Peninsula (Navarrete et al. 2017, 2023).
Secondly, variability δ13C and δ15N values could be due to foddering strategies. δ15N values could reflect the consumption of agricultural by-products such as crop stubble and other plants used as fodder. Moreover, the increase in 15N is not homogenous in plants, especially with manured cereal straws potentially being depleted in 15N compared to grains and legumes (Bogaard et al. 2007, 2013). The documented presence of collected bushes and branches such as Pistacia sp. and heather inside the cave (Buxó 1997) suggests that these resources might have complemented the diet of herds. Additionally, evidence of domestic activities within the cave, including processing and preparation of cereals and legumes for human consumption (Égüez et al. 2016; Martín-Socas et al. 2004) supports the hypothesis that the agricultural by-products were provided to the flocks. The neonate and infantile individuals recovered in the El Toro cave (30% of total of domestic caprines) suggest that animal foddering practices likely played a crucial role in maintaining the nutritional health of sheep mothers, vital for the survival of the lambs (Mellor and Stafford 2004).
Thirdly, the 15N-enriched values could result from confined feeding environment, such as a stable. This enrichment may occur due to the use of fodder and the presence of manure. Livestock manure is typically 15N-enriched compared to fodder (Bogaard et al. 2007; Commisso and Nelson 2006). Continuous use of the area where animals are housed can potentially enrich the soil with 15N (Commisso and Nelson 2006). In this sense, micromorphological analyses at El Toro cave indicate caprine stabling practices inside the cave (Égüez et al. 2016).
Pig management strategies
Pigs are likely omnivores and opportunistic feeders (Macdonald and Barrett 1993; Schley and Roper 2003). As omnivores, in the composition of collagen pig bone, nitrogen isotopes can come from both animal and plant proteins. Traditional pig husbandry may have involved home-based systems with complete to partial stabling near settlements or extensive herd management in semi-free to free-range regimes. This practice has been observed in modern traditional communities in northern Mediterranean (Albarella et al. 2007; Hadjikoumis 2012) and is also documented for prehistoric groups in Europe (Balasse et al. 2016) and in Northeast and Central Pyrenees of the Iberian Peninsula (Navarrete et al 2017; Villalba-Mouco et al. 2018).
At El Toro cave, the amplitude of variation for δ13C values of pigs is similar to that of wild herbivores (1.73‰), and their absolute δ13C values generally overlap with the herbivore data. There are no significant difference in average δ15N values between phases (Phase IV, 4.63‰; Phase IIIB, 4.05‰), and the 15N-enriched values (Phase IV, 3.26‰; Phase IIIB, 3.40‰) do not comprise the variation range of strict and wild herbivores at the site (2.8‰) (Figs. 3 and 4).
The diversity in 15N-enriched values indicates fluctuations in local livestock management practices (Navarrete et al. 2017). This suggests the pig diet may have mainly consisted of plant products, animal protein and agricultural by-products (cereals, pulses). Two management strategies align with these conditions. One involves household-level regimes or enclosed pigs. These pigs would mainly feed on plants (ground cereals and legumes), potentially supplemented with domestic leftovers, including animal products. This could result in higher δ15N values in pigs than local herbivores, though also depending on the quality and proportion of protein in their diets (Balasse et al. 2016; Madgwick et al. 2012; Müldner and Richards 2005). The high δ15N values (~ 6.5–-7.6‰) in some pig specimens could indicate some degree of manuring effect, which could then transfer to pig collagen through consuming animal products and waste. High values of δ15N are also documented in in pigs from Can Sadurní cave and Reina Amàlia-Caserna de Sant Pau sites (northeast Iberian Peninsula) during the Neolithic (Navarrete et al. 2017).
Second, pigs reared in semi-free or free-range systems would obtain most of their nutrients from available plants, although this does not exclude some consumption of small animals or insects and worms, as well as human food waste (Hamilton and Thomas 2012). Within this management strategy, we might expect pig collagen δ15N values to be similar or close to local herbivores (Madgwick et al. 2012, 2013). Similar interpretations have been proposed for prehistoric pigs possibly raised at the household-level with a mostly herbivorous diet (Balasse et al. 2016) and for modern pigs raised in traditional farming communities (Hadjikoumis 2012). Plant-based diets have been documented in the northeast Iberian Peninsula in sites of El Frare cave, La Draga and Serra de Mas Bonet (Navarrete et al. 2017) and in Central Pyrenees (Villalba-Mouco et al. 2018).
The pigs in El Toro cave present high variability in δ15N values, but this is not directly related to size. While there is considerable variability in pig size in the Iberian Peninsula (Navarrete and Saña 2017), the pigs from El Toro cave have a size associated with the domestic form according to the standardized criteria published by von den Driesch (1976) and Payne and Bull (1988). In addition, mortality profiles that these animals were mostly slaughtered between 14 and 21 months old and between 36 and 42 months old, when their reproductive capacity begins to decline. This mortality pattern has been documented in extensive pig-rearing livestock models of ethnographic societies (Hadjikoumis 2012). In extensive regimes, pigs are slaughtered at around 12 and 24 months old, while in intensive regimes, they are slaughtered younger, a consequence of faster fattening due to their strictly controlled diet (Hadjikoumis 2012). Even though the cave was seasonally occupied, during Phase IV and more intensively in Phase IIIB, and caprine penning inside the cave has been documented, there is no evidence of this practice in the case of pigs (Égüez et al. 2016). In this sense, the variability of isotopic data and archaeozoological data could indicate several feeding strategies for pigs.
Conclusion
Husbandry practices played an essential role in the economic strategies of El Toro cave during the Neolithic. Domestic species show a high percentage of representation compared to wild species. Among domesticates, caprines were most common, followed by pigs and cattle. Slaughtering patterns suggest meat was the focus of animal exploitation during Phase IV and Phase IIIB, especially for pigs and cattle. For caprines, meat-producing specimens dominated during Phase IV and Phase IIIB, although a significant increase in neonatal and infantile remains was recorded in Phase IIIB. This could suggest caprine birth in the cave, and the occurrence of slaughtering in the first 2 months is often associated with milk production (Vigne and Helmer 2007). Phase IIIB coincided with an intensification of agricultural practices with the synchronic use of the cave of humans and animals cohabitation (Martín-Socas et al. 1999, 2004).
Isotopic results from domesticates exhibit an extensive range of δ15N and δ13C values and provide evidence for diverse husbandry strategies and foddering regimes in El Toro cave during the Neolithic. The wide-ranging herbivore δ15N values from Phase IV and Phase IIIB suggest grazing on vegetation with a variety of δ15N values, potentially reflecting different manuring degrees on the fields. The isotopic heterogeneity in cattle suggests that the individuals are likely to come from disparate areas rather than just the immediate locality. Pigs were subjected to various foddering regimes, with high δ15N that may indicate a diet with animal protein and/or agricultural by-products. In addition, the age of killed pigs suggest a possible extensive regime. Homogeneity in their δ13C, similar to wild herbivores, may indicate their management at the local level.
In sum, this study provides new insights into Neolithic husbandry practices and presents new perspectives for analysing animal foddering strategies in mountain areas. In the future, the integration of these results with analyses of oxygen and strontium isotopes will help us understand the nature of mobility and the provenance of domesticates.
Data availability
All relevant data generated or analysed for this study are included in this published article.
Code availability
Not applicable
References
Acosta P (1995) Las culturas del Neolítico y Calcolítico en Andalucía occidental. Espacio, Tiempo y Forma. Serie I. Prehist Arqueol 8:33–80
Acosta P, Pellicer M (1990) La Cueva de la Dehesilla (Jerez de la Frontera): las primeras civilizaciones productoras en Andalucía Occidental. Consejo Superior de Investigaciones Científicas, Confederación Española de Centros de Estudios Locales, Jerez
Acosta P (1987) El Neolítico antiguo en el suroeste español. La Cueva de la Dehesilla (Cádiz). In: Guilaine J, Courtin J, Roudil J, Vernet J (eds) Premières Communautés Paysannes en Méditerranée occidentale. Actes du Colloque International du CNRS (Montpellier, 1983). Éditions du CNRS, Paris, pp 653- 659
Albarella U, Manconi F, Vigne JD, Rowley-Conwy P (2007) Ethnoarchaeology of pig husbandry in Sardinia and Corsica. In Albarella U, Dobney K, Ervynck A, Rowley-Conwy P (eds) Pigs and Humans. 10,000 Years of Interactions. Oxford University Press, Oxford, pp 285-307
Altuna J, Mariezkurrena K (2009) Tipos de cabañas ganaderas durante el Neolítico del País Vasco y zonas próximas. Archaeofauna 18:137–157
Ambrose SH (1990) Preparation and characterization of bone and tooth collagen for isotopic analysis. J Archaeol Sci 17:431–451. https://doi.org/10.1016/03054403(90)90007-R
Ambrose SH, Norr L (1993) Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate. Prehistoric human bone-archaeology at the molecular level. In: Lambert JB, Grupe G (eds) Prehistoric Human Bone. Springer, Berlin, pp 1–37. https://doi.org/10.1007/978-3-662-02894-0_1
Amorosi T (1989) A postcranial guide to domestic neo-natal and juvenile mammals: the identification and ageing of old world species (No. 533-534). British Archaeological Reports International Series 1533, Archaeopress, Oxford
Aura J E, Badal E, Borja G, García-Puchol O, Benito JL, Perez G, Perez-Ripoll M, Jordá JF (2005) Cueva de Nerja (Málaga). Los niveles neolíticos de la Sala del Vestíbulo. In: (coord.) Actas del III congreso del Neolítico en la Península Ibérica (Santander 5 a 8 de octubre de 2003). Santander, pp 975–987
Balasse M, Balasescu A, Tornero C, Frémondeau D, Hovsepyan R, Gillis R, Popovici D (2015) Investigating the scale of herding in chalcolithic pastoral communities settled by the Danube river in the 5th millennium BC: a case study at Bordusani-Popina and Hârsova-tell (Romania). Quat Int 436:29–40. https://doi.org/10.1016/j.quaint.2015.07.030
Balasse M, Evin A, Tornero C, Radu V, Fiorillo D, Popovici D, Andreescu R, Dobney K, Cucchi T, Bălăşescu A (2016) Wild, domestic and feral? Investigating the status of suids in the Romanian Gumelniţa (5th mil. cal BC) with biogeochemistry and geometric morphometrics. J Anthropol Archaeol 42:27–36. https://doi.org/10.1016/j.jaa.2016.02.002
Barone R (1976) Anatomie comparée des mammifères domestiques. Ostéologie 1:179–185
Boessneck J (1980) Diferencias osteológicas entre las ovejas (Ovis aries Linné) y cabras (Capra hircus Linné). In: Brothwell D, Higgs E (eds) Ciencia en arqueología. Fondo de Cultura Económica. México, pp 338–366
Bogaard A, Heaton TH, Poulton P, Merbach I (2007) The impact of manuring on nitrogen isotope ratios in cereals: archaeological implications for the reconstruction of diet and crop management practices. J Archaeol Sci 34:335–343. https://doi.org/10.1016/j.jas.2006.04.009
Bogaard A, Fraser R, Heaton TH, Wallace M, Vaiglova P, Charles M, Jones G, Evershed R, Stryring A, Andersen N, Arbogast RM, Bartosiewicz L, Gardeisen A, Kanstrup M, Maier U, Marinova E, Ninov L, Schäfer M, Stephan E (2013) Crop manuring and intensive land management by Europe’s first farmers. Proc Natl Acad Sci 110:12589–12594
Brown TA, Nelson EE, Vogel SJ, Southon JR (1988) Improved collagen extraction by modified Longin method. Radiocarbon 30:171–177. https://doi.org/10.1017/S0033822200044118
Buxó R (1997) Arqueología de las plantas. Crítica, Barcelona
Caceres I (2002) Estudio de los restos óseos de la fauna terrestre en el asentamiento de “El Retamar”. In: Ramos J, Lazarich M (eds) El asentamiento de “El Retamar” (Puerto Real). Contribucion al estudio de la Formacion Social Tribal y los inicios de la economía de produccion en la Bahía de Cadiz. Servicio de Publicaciones Universidad de Cadiz, Cadiz, pp 175–192
Camalich D, Martín-Socas D (2013) Los inicios del Neolítico en Andalucía. Tradición e Innovación. Menga. Rev Prehist Andalucía 4:103–129
Castaños P (2004) Estudio arqueozoológico de los macromamíferos del neolítico de la cueva de Chaves (Huesca). Saldvie 4:125–171
Commisso RG, Nelson DE (2006) Modern plant δ15N values reflect ancient human activity. J Archaeol Sci 33(8):1167–1176
Cuenca-Solana D, Rodríguez-Santos FJ, Nonza-Micaelli A, Cantillo-Duarte JJ, Clop-García X, Camalich-Massieu MD, Martín-Socas D (2021) Los instrumentos de concha y la producción alfarera durante el neolítico en la Cueva de el Toro (Antequera, Málaga). Monografies de la Societat ďHistòria Natural de les Balears p 32
DeNiro MJ, Epstein S (1978) Influence of diet on the distribution of carbon isotopes in animals. Geochim Cosmochim Acta 42:495–506
DeNiro MJ (1985) Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317:806–809. https://doi.org/10.1038/317806a0
Driesch A (1976) A guide to the measurement of animal bones from archaeological sites (Peabody Museum Bulletin 1). Peabody Museum of Archaeology and Ethnology, Cambridge
Égüez N, Mallol C, Martín-Socas D, Camalich MD (2016) Radiometric dates and micromorphological evidence for synchronous domestic activity and sheep penning in a Neolithic cave: Cueva de El Toro (Málaga, Antequera, Spain). Archaeol Anthropol Sci 8:107–123. https://doi.org/10.1007/s12520-014-0217-0
Fernàndez H (2001) Ostéologie comparée des petits ruminants eurasiatiques sauvages et domestiques (genres Rupicapra, Ovis, Capra et Capreolus): diagnose différentielle du squelette appendiculaire. Doctoral dissertation, University of Geneva
Filella I, Peñuelas J (2003) Partitioning of water and nitrogen in co-occurring Mediterranean woody shrub species of different evolutionary history. Oecologia 137:51–61. https://doi.org/10.1007/s00442-003-1333-1
Fraser RA, Bogaard A, Heaton T, Charles M, Jones G, Christensen BT, Halstead P, Merbach I, Poulton P, Sparkes D, Styring AK (2011) Manuring and stable nitrogen isotope ratios in cereals and pulses: towards a new archaeobotanical approach to the inference of land use and dietary practices. J Archaeol Sci 38:2790–2804. https://doi.org/10.1016/j.jas.2011.06.024
Friedli H, Lötscher H, Oeschger H, Siegenthaler U, Stauffer B (1986) Ice core record of the 13C/12C ratio of the atmospheric CO2 in the past two centuries. Nature 324:237–238. https://doi.org/10.1038/324237a0
Fuller BT, De Cupere B, Marinova E, Van Neer W, Waelkens M, Richards MP (2012) Isotopic reconstruction of human diet and animal husbandry practices during the Classical-Hellenistic, imperial, and Byzantine periods at Sagalassos, Turkey. Am J Phys Anthropol 149:157–171. https://doi.org/10.1002/ajpa.22100
García L, Wheatley DW (2009) El marco territorial de los dólmenes de Antequera: valoración preliminar de las primeras investigaciones. In: Ruiz B (ed) Dólmenes de Antequera. Tutela y valorización hoy. PH Cuadernos 23. Consejería de Cultura Junta de Andalucía, Sevilla, pp 128–143
Gillis R, Chaix L, Vigne JD (2011) An assessment of morphological criteria for discriminating sheep and goat mandibles on a large prehistoric archaeological assemblage (Kerma, Sudan). J Archaeol Sci 38(9):2324–2339. https://doi.org/10.1016/j.jas.2011.04.012
Grant A (1982) The use of tooth wear as a guide to the age of domestic animals. In: Wilson B, Grigson C, Payne S (eds.) Ageing and sexing animal bones from archaeological sites. British Archaeological Reports. International Series 1109, Archaeopress, Oxford, pp 91–108
Hadjikoumis A (2012) Traditional pig herding practices in Southwest Iberia: questions of scale and zooarchaeological implications. J Anthropol Archaeol 31(3):353–364. https://doi.org/10.1016/j.jaa.2012.02.002
Halley DJ, Rosvold J (2014) Stable isotope analysis and variation in medieval domestic pig husbandry practices in northwest Europe: absence of evidence for a purely herbivorous diet. J Archaeol Sci 49(1):1–5. https://doi.org/10.1016/j.jas.2014.04.006
Halstead P, Collins P (2002) Sorting the sheep from the goats: morphological distinctions between the mandibles and mandibular teeth of adult Ovis and Capra. J Archaeol Sci 29(5):545–553. https://doi.org/10.1006/jasc.2001.0777
Hamilton J, Thomas R (2012) Pannage, pulses and pigs: isotopic and zooarchaeological evidence for changing pig management practices in later Medieval England. Mediev Archaeol 56:234–259. https://doi.org/10.1179/0076609712Z.0000000008
Hammer Ø, Harper D, Ryan PD (2001) PAST: Paleontological Statistics Software Package for education and data analysis. Palaeontol Electron 4(1–4):1–9
Hammond C, O’Connor T (2013) Pig diet in medieval York: carbon and nitrogen stable isotopes. Archaeol Anthropol Sci 5:123–127. https://doi.org/10.1007/s12520-013-0123-x
Hellevang H, Aagaard P (2015) Constraints on natural global atmospheric CO2 fluxes from 1860 to 2010 using a simplified explicit forward model. Sci Rep 5:17352. https://doi.org/10.1038/srep17352
Jones GG (2006) Tooth eruption and wear observed in live sheep from Butser Hill, the Cotswold Farm Park and five farms in the Pentland Hills, UK. In: Ruscillo D (ed) Recent advances in ageing and sexing animal bones. Proceedings of the 9th Icaz Conference, Durham 2002. Oxbow Books, Oxford, pp 155–178
Lemoine X, Zeder MA, Bishop K, Rufolo S (2014) A new system for computing dentition-based age profiles in Sus scrofa. J Archaeol Sci 47(1):179–193. https://doi.org/10.1016/j.jas.2014.04.002
Lizcano R (1999) El Polideportivo de Martos (Jaén): un yacimiento neolítico del IV milenio A.C. Nuevos datos para la reconstrucción del proceso histórico del Alto Guadalquivir. Publicaciones CajaSur, Obra Social y Cultural. Córdoba
Longin R (1971) New method of collagen extraction for radiocarbon dating. Nature 230(5291):241–242
Lopez-Saez JA, Perez Díaz S, Alba Sanchez F (2011) Antropizacion y agricultura en el neolítico de Andalucía occidental a partir de la palinología. Menga. Revista de Prehistoria de Andalucía 2:73–85
Macdonald DW, Barrett P (1993) Mammals of Britain & Europe. HarperCollins, London, pp 312
Madgwick R, Mulville J, Stevens RE (2012) Diversity in foddering strategy and herd management in late Bronze Age Britain: an isotopic investigation of pigs and other fauna from two midden sites. Environ Archaeol 17(2):126–140. https://doi.org/10.1179/1461410312Z.00000000011
Madgwick R, Forest V, Beglane F (2013) Syndactyly in pigs: a review of previous research and the presentation of eight archaeological specimens. Int J Osteoarchaeol 23(4):395–409. https://doi.org/10.1002/oa.1260
Madgwick R, Esposito C, Lamb AL (2023) Farming and feasting at the Bronze Age-Iron Age transition in Britain (c. 900-500 BCE): multi isotope evidence for societal change. Front Environ Archaeol 2:1221581
Makarewicz CA (2014) Winter pasturing practices and variable fodder provisioning detected in nitrogen (δ15N) and carbon (δ13C) isotopes in sheep dentinal collagen. J Archaeol Sci 41:502–510
Makarewicz CA (2023) Extensive woodland pasturing supported Pitted Ware Complex livestock management systems: multi-stable isotope evidence from a Neolithic interaction zone. J Archaeol Sci 158:105689
Martín-Socas D, Massieu MDC, Herrero JLC, Rodríguez-Santos FJ (2018) The beginning of the Neolithic in Andalusia. Quat Int 470:451–471. https://doi.org/10.1016/j.quaint.2017.06.057
Martín-Socas D, Camalich MD, González P (1998) L’Andalusie (Espagne). Atlas du Néolithique Européen. 2-B, Études et Recherches Archéologiques de l’Université de Liège. Belgium, pp 871–933
Martín-Socas, D, Buxó R, Camalich MD, Goñi A (1999) Estrategias subsistenciales en Andalucía Oriental durante el Neolítico. SAGVNTVM Extra 2:25-30
Martín-Socas D, Camalich MD, González P (2004) La Cueva de El Toro (Sierra de El Torcal-Antequera-Málaga). Un modelo de ocupación ganadera en el territorio andaluz entre el VI y II milenios A.N.E. Arqueología Monografías. Consejería de Cultura Junta de Andalucía, Sevilla
Mellor DJ, Stafford KJ (2004) Animal welfare implications of neonatal mortality and morbidity in farm animals. Vet J 168(2):118–133
Minagawa M, Matsui A, Ishiguro N (2005) Patterns of prehistoric boar Sus scrofa domestication, and interisland pig trading across the East China Sea, as determined by carbon and nitrogen isotope analysis. Chem Geol 218(1–2):91–102. https://doi.org/10.1016/j.chemgeo.2005.01.019
Morales A, Martín JM (1995) Los mamíferos de la Cueva de Nerja (Analisis de las cuadrículas NM-80A NM-80B y NT-82). In: Pellicer M, Morales A (eds) Fauna de la Cueva de Nerja I. Salas de la Mina y de la Torca, Campañas 1980-82: 59-159. Trabajos sobre la Cueva de Nerja 5
Morales A. Riquelme JA (2004) Faunas de mamíferos del Neolítico andaluz: tendencias diacrónicas fiables. Actas del II y III Simposio de Prehistoria Cueva de Nerja. Fundación Cueva de nerja, España, pp 41–51
Müldner G, Richards MP (2005) Fast or feast: reconstructing diet in later medieval England by stable isotope analysis. J Archaeol Sci 32(1):39–48. https://doi.org/10.1016/j.jas.2004.05.007
Navarrete V, Saña M (2017) Size changes in wild and domestic pig populations between 10,000 and 800 cal. BC in the Iberian Peninsula: evaluation of natural versus social impacts in animal populations during the first domestication stages. Holocene 27(10):1526–1539. https://doi.org/10.1177/0959683617693902
Navarrete V, Colonese AC, Tornero C, Antolín F, Von Tersch M, Subirà E, Saña M (2017) Feeding management strategies among the early Neolithic pigs in the NE of the Iberian Peninsula. Int J Osteoarchaeol 27(5):839–852. https://doi.org/10.1002/oa.2598
Navarrete V, Tornero C, Balasse M, Saña M (2019) Food management of early introduced caprine and bovine herds in the early Neolithic site of La Draga (Banyoles): an isotopic approach. Int J Osteoarchaeol 29:986–998. https://doi.org/10.1002/oa.2812
Navarrete V, Viñerta A, Clemente-Conte I, Gassiot E, Rey Lanaspa J, Saña M (2023) Early husbandry practices in highland areas during the Neolithic: the case of Coro Trasito cave (Huesca, Spain). Front Environ Archaeol 2:1309907. https://doi.org/10.3389/fearc.2023.1309907
O’Leary MH (1981) Carbon isotope fractionation in plants. Phytochemistry 20(4):553–567
O’Leary MH (1988) Carbon isotopes in photosynthesis: fractionation techniques may reveal new aspects of carbon dynamics in plants. Bioscience 38:328–336
Oueslati T (2006) Approche archéozoologique des modes d’acquisition, de transformation et de consommation des ressources animales dans le contexte urbain gallo-romain de Lutèce (Paris, France). Series de British Archaeological Reports 1479, Oxford
Payne S (1985) Morphological distinctions between the mandibular teeth of young sheep, Ovis, and goats, Capra. J Archaeol Sci 12(2):139–147
Payne S (1987) Reference codes for wear states in the mandibular cheek teeth of sheep and goats. J Archaeol Sci 14(6):609–614
Payne S, Bull G (1988) Components of variation in measurements of pig bones and teeth, and the use of measurements to distinguish wild from domestic pig remains. ArchæoZoologia 2:27–65
Pechenkina EA, Ambrose SH, Xiaolin M, Benfer RA (2005) Reconstructing northern Chinese Neolithic subsistence practices by isotopic analysis. J Archaeol Sci 32(8):1176–1189. https://doi.org/10.1016/j.jas.2005.02.015
Pellicer M, Acosta P (1982) El Neolítico Antiguo en Andalucía Occidental. Le Neolithique Ancien Mediterraneen. Actes du Colloque Internacional de Prehistoire (Montpellier 1981), Archeologie en Languedoc, Montpellier, pp 49-60
Peña-Chocarro L, Perez Jorda G, Morales Mateos J, Vera Rodríguez JC (2013) Y llegaron los agricultores: agricultura y recoleccion en el occidente del Mediterraneo. In: Martín-Socas D, Camalich MD (eds) Dossier: Entre Dos Aguas. Tradicion e Innovacion en las Sociedades Neolíticas de Andalucía, Menga 4:15–33
Perdiguero M (1981) Materiales cerámicos neolíticos de la Sierra del Torcal, Antequera, Málaga. Mainake II-III:30–47
Pérez-Ripoll M (2013) Una visión de la arqueozoología valenciana a través del tiempo. Animals i arqueologia hui, 3
Pérez-Ripoll M (2016) Los niveles de corral en el yacimiento neolítico de la Cova de l’Or, sectores K34, K35 y K36. Información aportada por el estudio de la fauna. Del Neolític a l'Edat de Bronze en el Meditarrani occidental: estudis en homenatge a Bernat Martí Oliver. Museu de Prehistòria de València, pp 117–124
Prummel W, Frisch HJ (1986) A guide for the distinction of species, sex and body size in bones of sheep and goat. J Archaeol Sci 13:567–577
Riquelme JA (1998) Contribución al Estudio Arqueofaunístico durante el Neolítico y la Edad del Cobre en las Cordilleras Béticas: el Yacimiento Arqueológico de Los Castillejos en Las Peñas de los Gitanos, Montefrío (Granada). Doctoral Dissertation, Universidad de Granada
Rodríguez-Ariza O (1996) Análisis antracológicos de yacimientos Neolíticos de Andalucía. Actas del 1 Congreso del Neolític a la Península Iberica. Formació i implantació de les comunitats agrícoles (Gavá-Bellaterra 1995). Rubricatum 1 (1):73–83
Rodríguez-Ariza O (2004) Estudio antracologico. In: Martín-Socas D, Camalich Massieu MD, González Quintero P (eds) La Cueva de El Toro (Sierra de El Torcal-Antequera-Malaga). Un modelo de Ocupacion Ganadera en el Territorio Andaluz entre el VI y II Milenios A.N.E. Arqueología Monografías 21, Junta de Andalucía, Sevilla, pp 61–79
Rodríguez-Rodríguez A, Martín-Socas D, Cámallich Massieu MD, González Quintero P (1996) Las actividades tecnoeconómicas en “Cueva del Toro” (Antequera, Málaga) a través del análisis funcional. Actas del I Congrés del Neolític a la Península Iberica. Formació i implantació de les comunitats agrícoles (Gava-Bellatera 1995), Rubricatlllll 1 (l):161–165
Rodríguez-Rodríguez AC (2004) Análisis funcional de los instrumentos líticos tallados. In, La Cueva de El Toro (Sierra de El Torcal, Antequera- Málaga).Un modelo de ocupación ganadera en territorio andaluz: entre el VI y II milenios A.N.E., (dir., Martín Socas, D., M.D. Camalich Massiey y P. González Quintero), Arqueología Monografías. Junta de Andalucía, Consejería de Cultura, Sevilla, pp 35–160
Rovira, N (2007) Agricultura y gestión de los recursos vegetales en el sureste de la Península Ibérica durante la prehistoria reciente. Doctoral Dissertation, Universidad Pompeu Fabra
Salvagno L, Albarella U (2017) A morphometric system to distinguish sheep and goat postcranial bones. PloS one 12(6):e0178543. https://doi.org/10.1371/journal.pone.0178543
Salvatierra V (1994) Historia y desarrollo del Modelo Andaluz de Arqueología. Trab Prehist 51:1–13
Saña M (2013) Domestication of animals in the Iberian Peninsula. In: Colledge S, Conolly J, Dobney K, Manning K, Shennan S (eds) The origins and spread of domestic animals in southwest Asia and Europe. UCL Institute of Archaeology Publications, London, pp 195–220
Saña M, Antolín F, Alcántara R, Sierra A, Tornero C (2020) Integrating domesticates: earliest farming experiences in the Iberian Peninsula. Farmers as the frontier: a pan European perspective on Neolithisation, pp 161–176
Sanchidrián JL, García M (1987) Material cerámico de la Sima Hoyo del Tambor (El Torcal, Antequera). Mainake 8–9:89–104
Santana J, Rodríguez-Santos FJ, Camalich-Massieu MD, Martín-Socas D, Fregel R (2019) Aggressive or funerary cannibalism? Skull-cup and human bone manipulation in Cueva de El Toro (Early Neolithic, southern Iberia). Am J Phys Anthropol 169(1):31–54. https://doi.org/10.1002/ajpa.23805
Schley L, Roper TJ (2003) Diet of wild boar Sus scrofa in Western Europe, with particular reference to the consumption of agricultural crops. Mamm Rev 33:43–56. https://doi.org/10.1046/j.1365-2907.2003.00010.x
Silver IA (1969) The ageing of domestic animals. In: Brothwell R, Higgs ES (eds) Science in archaeology: a survey of progress and research. Thames & Hudson, London, pp 283–302
Szpak P (2014) Complexities of nitrogen isotope biogeochemistry in plant–soil systems: implications for the study of ancient agricultural and animal management practices. Front Plant Sci 5:288. https://doi.org/10.3389/fpls.2014.00288
Tarifa-Mateo N, Clop-García X, Rosell-Melé N,Camalich-Massieu MD, Comes-Bordas P, Martín-Socas D, Nonza-Micaelli A, Rodríguez-Santos FJ (2019).New insights from Neolithic pottery analyses reveal subsistence practices and pottery use in early farmers from Cueva de El Toro (Málaga, Spain). Archaeol Anthropol Sci. https://doi.org/10.1007/s12520-019-00874-9
Tieszen LL (1991) Natural variations in the carbon isotope values of plants: implications for archaeology, ecology, and paleoecology. J Archaeol Sci 18:227–248
Vaiglova P, Bogaard A, Collins M, Cavanagh W, Mee C, Renard J et al (2014) An integrated stable isotope study of plants and animals from Kouphovouno, southern Greece: a new look at Neolithic farming. J Archaeol Sci 42:201–215. https://doi.org/10.1016/j.jas.2013.10.023
Valente MJ (2016) Zooarqueologia do Neolítico do Sul de portugal: Passado, presente e futuros. In: Diniz M, Neves C, Andrea M (eds) O Neolítico em Portugal antes do horizonte 2020: perspectivas em debate. AAP, Lisboa, pp 87–108
Van Klinken GJ (1999) Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J Archaeol Sci 26:687–695. https://doi.org/10.1006/jasc.1998.0385
Vigne JD, Helmer D (2007) Was milk a “secondary product” in the Old World Neolithisation process? Its role in the domestication of cattle, sheep and goats. Anthropozoologica 42(2):9–40
Villalba-Mouco V, Utrilla P, Laborda R, Lorenzo JI, Martínez-Labarga C, Salazar-García DC (2018) Reconstruction of human subsistence and husbandry strategies from the Iberian Early Neolithic: a stable isotope approach. Am J Phys Anthropol 167(2):257–271. https://doi.org/10.1002/ajpa.23622
Watson J, Paz MA, Tusell M, Mañosa M (2004) Análisis arqueofaunístico. Campaña de 1988. In: Martín-Socas D, Camalich MD, González Quintero P (eds) La Cueva de El Toro (Sierra de El Torcal, Antequera- Málaga).Un modelo de ocupación ganadera en territorio andaluz: entre el VI y II milenios A.N.E. Arqueología Monografías. Junta de Andalucía, Sevilla, pp 215-266
Yousfi S, Serret MD, Voltas J, Araus JL (2010) Effect of salinity and water stress during the reproductive stage on growth, ion concentrations, Δ13C, and δ15N of durum wheat and related amphiploids. J Exp Bot 61:3529–3542
Zapata L, Peña Chocarro L, Perez Jorda G, Stika HP (2005) Difusion de la agricultura en la Península Iberica. In: Arias Cabal P, Ontañon Peredo R, García-Monco C (eds) Actas del III Congreso de Neolítico en la Península Iberica (Santander 2003), Santander, pp 103–113
Zavodny E, McClure SB, Culleton BJ, Podrug E, Kennett DJ (2014) Neolithic animal management practices and stable isotope studies in the Adriatic. Environ Archaeol 19(3):184–195
Zeder MA (2006) Central questions in the domestication of plants and animals. Evol Anthropol 15(3):105–117. https://doi.org/10.1002/evan.20101
Zeder MA, Lapham HA (2010) Assessing the reliability of criteria used to identify postcranial bones in sheep, Ovis, and goats. Capra. J Archaeol Sci 37(11):2887–2905. https://doi.org/10.1016/j.jas.2010.06.032
Zeder MA, Pilaar SE (2010) Assessing the reliability of criteria used to identify mandibles and mandibular teeth in sheep, Ovis, and goats, Capra. J Archaeol Sci 37:225–242. https://doi.org/10.1016/j.jas.2009.10.002
Acknowledgements
The authors thank the anonymous reviewers for their valuable comments on this article. The authors thank Dr. Pau Comes (ICTA-UAB) for his help with the IRMS data and Dr. Eulalia Subirá (UAB) for her help with collagen extraction. Research by VN was supported by the research project CEECIND/03351/2020 (Fundação para a Ciência e a Tecnologia, Portugal). RA is a Margarita Salas (Spanish Ministry of Universities and European Union-NextGenerationEU programme) postdoctoral researcher. AS is a JdC-Formación postdoctoral researcher. Part of this research has been carried out in 2021 SGR 00527 (EarlyFoods).
Funding
Open access funding provided by FCT|FCCN (b-on). The research was supported by the Ministerio de Economía y Competitividad, Spain, HAR2017-88304-P (MS), PID2019-104442GB-100 (MSC and DMS) and PID2020-115715GB-I00 (MS), and ICREA Academia programme by MS.
Author information
Authors and Affiliations
Contributions
V. Navarrete: conceptualization, data curation, investigation, methodology (faunal analysis and stable isotope analysis), writing-original draft, review and editing. A. Sierra: conceptualization, investigation, writing—review and editing. R. Alcàntara: conceptualization, investigation, methodology (faunal analysis), writing—review and editing. MD. Camalich: conceptualization, funding acquisition, investigation, writing—review and editing. D. Martín-Socas: conceptualization, funding acquisition, investigation, writing—review and editing. M. Saña: conceptualization, funding acquisition, investigation, methodology (faunal analysis), writing—review and editing.
Corresponding author
Ethics declarations
Ethics approval
Not applicable
Consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Navarrete, V., Sierra, A., Alcàntara, R. et al. Integrative approaches to the study of animal management practices during the Neolithic of South Iberian Peninsula: the case of El Toro cave (Antequera, Málaga, Spain). Archaeol Anthropol Sci 16, 21 (2024). https://doi.org/10.1007/s12520-023-01926-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-023-01926-x