
Abstract
Discerning vulnerability differences among different aged trees to drought-driven growth decline or to mortality is critical to implement age-specific countermeasures for forest management in water-limited areas. An important species for afforestation in dry environments of northern China, Mongolian pine (Pinus sylvestris var. mongolica Litv.) has recently exhibited growth decline and dieback on many sites, particularly pronounced in old-growth plantations. However, changes in response to drought stress by this species with age as well as the underlying mechanisms are poorly understood. In this study, tree-ring data and remotely sensed vegetation data were combined to investigate variations in growth at individual tree and stand scales for young (9 − 13 years) and aging (35 − 52 years) plantations of Mongolian pine in a water-limited area of northern China. A recent decline in tree-ring width in the older plantation also had lower values in satellited-derived normalized difference vegetation indices and normalized difference water indices relative to the younger plantations. In addition, all measured growth-related metrics were strongly correlated with the self-calibrating Palmer drought severity index during the growing season in the older plantation. Sensitivity of growth to drought of the older plantation might be attributed to more severe hydraulic limitations, as reflected by their lower sapwood- and leaf-specific hydraulic conductivities. Our study presents a comprehensive view on changes of growth with age by integrating multiple methods and provides an explanation from the perspective of plant hydraulics for growth decline with age. The results indicate that old-growth Mongolian pine plantations in water-limited environments may face increased growth declines under the context of climate warming and drying.
Similar content being viewed by others


Introduction
Tree growth decline and mortality have recently occurred across numerous ecosystems globally which are likely associated with increases in intensity and frequency of droughts (Allen et al. 2010; IPCC 2013, 2021). Drought negatively affects growth and carbon sequestration capacity of forest ecosystems and hence can have a significant effect on the global carbon cycle (Ciais et al. 2005; Sippel et al. 2018). Endogenous processes related to age have been suggested as an important factor affecting variability of tree growth and its response to water availability (McMahon et al. 2010; Luo and Chen 2013; Chen et al. 2016). In the process of development, trees initially grow slowly, then have a period of increasing growth, followed by decreasing growth, all associated with changes of biological processes with aging (Fritts 1976). A number of studies have reported that the growth behavior of different aged trees reflected hydroclimate variability and this was linked to the changes in tree physiology with increasing age (Carrer and Urbinati 2004; Rozas et al. 2009; McDowell and Allen 2015). For example, evidence from natural forests in some high latitude areas indicated that old trees were more vulnerable to growth decline and mortality under drought stress than younger ones (Chen et al. 2016; Liu et al. 2021). In contrast, several dendroclimatological studies have shown that radial growth of younger trees is more likely to be affected by climate change relative to that of older trees (Carrer and Urbinati 2004; Rozas et al. 2009). A study investigating performance of a natural white spruce forest in western Canada found that recent climate drying led to growth decreases in both old and young trees (Hogg et al. 2017). However, few studies have examined changes in growth response to water availability across different ages in shelterbelts that are usually established in water deficit areas. Determining whether young or old trees are more susceptible to drought-related growth decline and underlying mechanisms is crucial to the construction and management of shelterbelts, as it can provide important information for making age-specific adaptation strategies.
Variations of tree growth performances under drought are closely associated with changes of xylem hydraulic behavior (Tyree 2003; Choat et al. 2018). For example, canopy conductance decreases under climate warming and drying through increasing vapour pressure deficits which would adversely affect photosynthesis and growth (Whitehead and Jarvis 1981; McDowell and Allen 2015). In addition, taller old-growth trees enduring greater resistance in water transport because of longer hydraulic paths faced higher risks of growth decline or mortality, compared with shorter, younger trees (Enquist 2003; Petit et al. 2011; McDowell and Allen 2015). Older, taller trees are thus considered more subject to growth decline or even mortality associated with severe hydraulic limitations, particularly given a drier future climate (Bennett et al. 2015). An increasing number of studies have reported growth decline and death linked to drought across different biomes. Large, old individuals were the main victims that influenced the sustainable development of forests as well as their capacity for carbon sequestration (Allen et al. 2010; Ryan 2015; Gora and Esquivel-Muelbert 2021). As for different aged shelterbelts in water-limited areas, investigating temporal changes of growth performance with age and clarifying underlying mechanisms from the perspective of plant hydraulics will provide important information for the management of mature forests vulnerable to water stress.
Tree-ring analysis provides an ideal approach to explore long-term variations in radial growth on an annual time scale in their response to environmental factors (Fritts 1976). Using dendroclimatology, the temporal variability of tree growth covering the entire life cycle can be revealed, which will be beneficial for characterizing differences of growth performances at various ages and identifying underlying causes (Fritts 1976; Schweingruber 1988). However, this method focuses on growth performance in the transverse direction of the trunk for individual trees and is often restricted to tree growth behavior in a limited number of sites (Schweingruber 1988; Cook and Kairiukstis 1990). Compared with tree-ring data, time series remote-sensing vegetation indices (VIs) based on spectral reflectance measurements of satellite images can be used to monitor temporal changes and spatial patterns of canopy conditions at the stand level as well as aboveground productivity at regional and even global levels (West et al. 2019; Liu et al. 2021). Among the VIs, the normalized difference vegetation index (NDVI) is most commonly used for examining the vegetation greenness at a canopy scale under environmental stress (Tucker 1979). In recent decades, NDVI has been used successfully to monitor drought impacts on vegetation growth at regional, continental, and global scales (Kogan 1990; Karnieli et al. 2010; Vicente-Serrano et al. 2013; Ukkola et al. 2021). The normalized difference water index (NDWI), another vegetation index, is sensitive to fluctuations of vegetation moisture condition and commonly serves as a surrogate of canopy water content (Gao 1996; Chen et al. 2005; Ji et al. 2011). The combination of NDVI and NDWI makes it possible to examine the relationship between vegetation growth status and water content that would help to reveal the underlying mechanisms behind the fluctuations of canopy vitality. Both tree-ring parameters and time series VIs have been widely used for investigating long-term growth and their response to climate variability, with focus respectively on individual tree and forest canopy scales. However, little research has attempted to combine these two methods (Kannenberg et al. 2019). Examining tree growth from different spatial scales by integrating multiple methods, i.e., tree-ring and remote-sensing analyses, would help to detect signs of tree growth decline and comprehensively assess forest dynamics for shelterbelts established in water-deficit areas.
Mongolian pine (Pinus sylvestris var. mongolica Litv.) is widely used for establishing shelterbelts in water-deficit areas of northern China (Zhang et al. 2005a). However, in these areas, the species is vulnerable to growth decline and mortality when it reaches 30 − 35 years old (below its lifespan of 150 − 200 years in its natural areas), a phenomenon called “premature aging” (Song et al. 2016; Zhu et al. 2016). A recent study attributed the decrease in radial growth in introduced areas to increasing water stress with aging and explained the mechanisms behind this phenomenon from the view of xylem hydraulics (Liu et al. 2018). However, less is known about the responses of growth to water availability at the stand level, as well as the response across different ages for this species, hindering a more comprehensive understanding on forest response to water stress with increasing stand age. In this study, both dendroclimatological analysis and remotely sensed Vis were used to investigate growth changes to drought in relation to changes in xylem hydraulics in two age groups (9 − 13 years and 35 − 52 years) of Mongolian pine plantations in a water-limited area of northern China. It was hypothesized that: (1) interannual variations in tree-rings and of remotely sensed VIs were coupled in both older and younger plantations; (2) both tree-ring values and remotely sensed VIs were more sensitive to drought in older plantations; and, (3) xylem hydraulic conductivities decreased in the older Mongolian pine trees.
Materials and methods
Description of study area and tree species
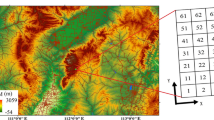
The study area is located in the southern part of the Horqin Sandy Land in northern China. This region has a temperate continental monsoon climate with a May to September growing season receiving 85% of the annual precipitation (Fig. S1 in Online Appendix). Annual total precipitation was 512 mm and mean temperature 7.6 °C during 1953 − 2020 according to climate records from the Zhangwu meteorological station (42.42°N, 122.53°E) from the China Meteorological Administration. The forest vegetation is composed of a number species used for afforestation such as Pinus sylvestris var. mongolica Litv., Pinus tabulaeformis Carr., Pinus densiflora Siebold & Zucc. var. zhangwuensis Zhang and Yuan, Populus simonii Carr., Populus alba 'Berolinensis' L., and several native species (e.g., Ulmus pumila L., Phellodendron amurense Rupr.) that are naturally distributed. Historically, this region has been alternately occupied by forest-steppes and deserts under the influences of both climate fluctuations and human activities (Feng and Jiang 1996). Under increasing drought, grazing and aggressive land reclamation, native forests and grasslands were gradually transformed into desert lands from the late nineteenth century to the early twentieth century (Tao 1987). Massive reforestation programs organized by the government since the 1950s have returned parts of the desert lands back to forest that effectively curtailed further extension of deserts and protected the ecological environment of urban settlements in surrounding areas (Zhu et al. 2016).
Mongolian pine is a variety of Scotch pine (Pinus sylvestris L.) in eastern Asia. Because of its ability in resisting drought and adapting to sandy soils, Mongolian pine was introduced from its native areas in Honghuaerji, Hulunbuir, China for afforestation to the region since the 1950s (Zhu et al. 2006). This species has also been widely used for afforestation programs in other areas of northern China over the past decades and have reached more than northern 300 counties (Zhu et al. 2005b), playing important roles in increasing vegetation cover and improving ecological services.
Data collection
Acquiring tree-ring data
During August 2020, two different age plantations of this species (9 − 13 years and 35 − 52 years) in the southern Horqin sandy lands in northern China. Age was obtained by counting the number of annual rings at breast height. Two plots of each age group were selected (42.71434 − 42.71479°N, 122.49038 − 122.49099°E; 42.66875 − 42.66920°N, 122.57166 − 122.57227°E), with one plot per plantation for annual ring sampling. Tree heights in the younger and older plantations were approximately 2.5 − 3 m and 13 − 14 m, respectively. Thirty-seven and 33 increment cores were taken from 20 to 21 trees, respectively, with one or two cores per tree at breast height using an increment borer. The increment cores were processed to obtain ring width data as well as their cross-dated results following the standard procedure of tree-ring analysis (Cook and Kairiukstis 1990). To extract the variances linked to climate while removing those related to biological trends, the tree-ring width measurements of each core for the older plantation were detrended using a cubic spline with a 50% frequency–response cutoff equal to two-thirds of the series length, which was performed by the program ARSTAN (Cook 1985). All detrended series were averaged to obtain a ring width chronology for the older plantation using the method of bi-weight robust means (Cook and Kairiukstis 1990; Fig. 1). For the younger plantation, the undetrended ring width series of all cores were averaged to obtain a raw chronology through the way of bi-weight robust means (Fig. 1a). We derived tree-ring raw chronology rather than the standard detrended chronology because the period of tree-ring data availability was relatively short (2008 − 2019). This is because there could be some possible drawbacks on detrending tree-ring data of short periods using the current detrending methods (Fritts 1976; Cook 1985). However, the raw tree-ring width (RRW) data of the younger plantation could represent those being detrended, since trends of biology related to age might have limited impacts on annual ring data covering a relatively short period (Cook and Kairiukstis 1990). Chronology statistics include mean sensitivity (MS), standard deviation (SD), inter-series correlation (Rbar) and expressed population signal (EPS; Fritts 1976; Table 1). For each chronology, the reliable period is determined when EPS surpasses 0.85 (Wigley et al. 1984). Accordingly, the reliable periods for the chronologies of the younger and older plantations were 2010 − 2019 and 1983 − 2019, respectively (Fig. 1a, b). Tree-ring chronologies covering these periods were used for growth-climate correlation analysis.

a Variability in annual ring width and width indexes over time respectively for the younger (PS1) and b older age groups (PS2) as well as changes of sample depth over time for the two age groups; c change of basal area increment (BAI) over time for the two age groups; sample depth denotes the number of increment cores (light blue area). The bold oblique line shows a significant decreasing (P < 0.05) trend in the tree-ring width index over the period of 2004 − 2019; downward arrow marks the time when the expressed population signal (EPS) of the tree-ring width raw chronology or tree-ring width standard chronology is larger than 0.85. The gray area denotes the common period for the BAI of the two groups
To better reflect radial growth of different aged Mongolian pine, annual basal area increment (BAI) was also calculated using non-detrended tree-ring data through the formula of Biondi and Qeadan (2008):
where BAIt is BAI in year t; RWt and RWt-1 are cumulative undetrended ring widths from the pith to year t and t-1, respectively.
Tree ages denote the cambium ages of the trees in 2020 when the tree-ring cores were sampled; the end of the chronology extension for both groups was determined in 2019, as the complete rings for the trees in 2020 had not formed when they were sampled in early August of that year. MS is mean sensitivity; SD is standard deviation; Rbar is inter-series correlation; EPS is expressed population signal. The unit of tree-ring width raw series was mm, which was used for calculating chronology statistics for PS1.
Acquiring remotely sensed vegetation index data
Six cloud-free sentinel-2A Level-1C MSI images (tile number T51TVH) covering the same region (including our study area) were acquired between late July and late August during 2016 − 2021, with one image per year (Table S1 in Online Appendix). The sentinel-2A satellite was launched by the European Space Agency 23rd June 2015 to provide high spatial resolution multispectral data covering the globe with a 5-day revisit frequency (Drusch et al. 2012). The Level-1C images used in this study were derived from the Sentinels Scientific Data Hub (https://scihub.copernicus.eu/) which had already been orthorectified and geometrically corrected (Korhonen et al. 2017). Therefore, radiometric calibration and atmospheric correction for the images were further carried out utilizing the Sen2cor 2.9.0 program (Gascon et al. 2017), through which Level-1C data were converted into Level-2A data. All 13 spectral bands with a spatial resolution between 10 and 60 m for each image were combined and converted into the spatial resolution of 10 m using the ArcGIS 10.2 software. The analyzed images were further extracted from the combined images using the vector boundary of the study area in the ERDAS 2010 software (ERDAS 1999).
After the above preprocessing, the spectral reflectance values from band 4 (red region), band 8 (near infrared region) and band 12 (short wave infrared region) of the images were employed using formulas (2) and (3) for obtaining NDVI and NDWI data, respectively, which were performed with the ERDAS 2010 software.
NDVI was calculated using the following formula (Tucker 1979):
where \(\rho_{{{\text{NIR}}}}\) and \(\rho_{{{\text{Re}} d}}\) are spectral reflectance values in near infrared (NIR) and red regions, respectively.
NDWI was calculated using the formula (Chen et al. 2005):
where \(\rho_{SWIR}\) is the value of spectral reflectance in the short wave infrared region (SWIR).
NDVI and NDWI data for the younger and older Mongolian pine plantations were obtained through the following steps. Five square plots in each of the two plantations were established according to the layouts of pixels in the sentinel-2A Level-1C MSI image, with one covering the tree-ring sampling plot as described previously and surrounded by the other four for remotely sensed analysis. Each plot covered 5 pixels × 5 pixels of the image corresponding to 50 m × 50 m in the field. The NDVI and NDWI data of each square plot were extracted from the data layers of these two indicators. The data for the five plots within each plantation were averaged and further used for analyzing canopy vegetation conditions at the stand scale.
Acquiring xylem hydraulic data
On early mornings during August 2020, four branches facing the sun, with per branch from each tree, were collected at the tree-ring sampling plot within each of the plantations for hydraulic conductivity measurements. During the sampling process, the cut ends of the four branches were recut immediately under water to eliminate potentially embolized tracheids. In the laboratory, 10-cm stem segments were extracted from the branches under water. The ends of each segment were then connected to a tubing apparatus filled with a degassed and filtered 20 mM KCl solution and a graduated glass pipette for measuring native hydraulic conductivity (Kh-native; Sperry and Tyree 1988). The solution flowed through the segment under a hydrostatic pressure developed by a hydraulic head of 50 cm. Native hydraulic conductivity (Kh-native, kg m s−1 MPa−1) was calculated using the following formula (Tyree and Ewers 1991):
where Jv denotes the flow rate through the stem segments (kg s−1); ΔP/ΔL is the pressure gradient across the stem segment (MPa m−1).
Sapwood-specific (Ks, kgm−1 s−1 MPa−1) and leaf-specific (Kl, kg m−1 s−1 MPa−1) hydraulic conductivities were obtained by Kh-native divided by the sapwood area (SA) of the transverse section and the total area of the needles terminal to the segment (LA), respectively. Both SA and LA were calculated using the software ImageJ 1.48v (US National Institutes of Health, Bethesda, MD, USA). The measurements of the two variables provided LA to SA ratio (LA/SA).
Statistical analysis
To investigate long-term growth responses to drought at both individual tree and stand scales, Pearson correlation analyses between the tree-ring metrics (e.g., RRW and tree-ring width index—RWI), BAI, NDVI and NDWI of the younger and older plantations and monthly self-calibrating Palmer drought severity index (scPDSI; Wells et al. 2004) were carried out during their overlapping period. Monthly scPDSI data were obtained by averaging the corresponding gridded data within the geographical range 42.5 − 43.0 N, 122 − 123 E from the CRU scPDSI Global 4.05early datasets with a spatial resolution of 0.5° × 0.5° (van der Schrier et al. 2013; Barichivich et al. 2020). This range covered all the sampling plots. The available data of scPDSI included the period from January 1901 to December 2020. The scPDSI data during January through October were used.
To test differences in the traits of xylem hydraulics and the remotely sensed vegetation indices between the younger and older age groups, a one-way analysis of variance (ANOVA) was carried out using the software SPSS 13.0 (SPSS Inc. 1997).
Results
Differences in growth patterns between younger and older age groups
There were large differences in radial growth patterns between the two age groups. The RRW of the younger group (PS1) showed an overall increase before 2012 but decreased afterwards (Fig. 1a), which may partly be attributed to the effects of biological trends. Inconsistently, RWI of the older group (PS2) exhibited a significant (P < 0.05) period of decline, i.e., 2004 − 2019, while there was no significant trend over the earlier period (Fig. 1b). Both Rbar and EPS for the RRW of PS1 and the RWI of PS2 were relatively high (Table 1), indicating that the radial growth of the two age groups might be sensitive to climate variability. During the common period (2008–2019) of the two age groups, the values of BAI for PS1 showed an obvious increase but PS2 remained relatively stable (Fig. 1c).
Consistent with the patterns of individual radial tree growth, vegetation greenness and water content at the stand level were poorest in the older plantations than in the younger ones. NDVI values of PS2 were lower than those of PS1 during the common period 2016 − 2021, with significant disparities between both groups in 2017, 2019 and 2021 (P < 0.1; Fig. 2a). Lower NDWI values were also observed for PS2 than in PS1 over the same years, with a significant difference in 2021 (P < 0.05; Fig. 2b).

Variability in remote sensing-based Normalized Difference Vegetation Index (NDVI; a and Normalized Difference Water Index (NDWI; b respectively for the younger (PS1) and older (PS2) groups of P. sylvestris var. mongolica during the peak of growing season (late July or late August) over the period of 2016 − 2021. Bar above/below each solid circle denotes 1 standard error (n = 5); * and ** respectively denote statistically significant difference at the level of 0.1 and 0.05 between the two age groups
There did not appear to be any close relationships between the metrics of radial growth at a single tree level and those at a stand level. Correlations of tree-ring metrics (RRW and RWI) and BAI with NDVI and NDWI during the common period 2016 − 2019 were all insignificant (P > 0.1) in each of the age groups (results not shown). However, it should be noted that there may be large uncertainties on the correlation results between these two kinds of indicators because the period of correlation analysis was relatively short, and hence their relationships may need to be further verified using longer time series data in future studies. There were strong associations between individual radial growth at the tree level and at the stand level in the same age group. Specifically, RWI was significantly (P < 0.01) and positively correlated with BAI in PS2, although there was no significant correlation in PS1 (Fig. 3a). Moreover, NDVI was significantly (P < 0.01) correlated with NDWI in both age groups (Fig. 3b).

Scatter plots showing the relationships of annual basal area increment (BAI) with tree-ring metrics and of normalized difference vegetation index (NDVI) with normalized difference water index (NDWI) over their common periods (2008 − 2019, 2016 − 2021, respectively), for the younger (PS1) and older (PS2) groups of P. sylvestris var. mongolica. The tree-ring metrics used for PS1 and PS2 are raw tree-ring width and tree-ring width index, respectively. The solid and dashed oblique lines respectively show significant (P < 0.05) and insignificant trends for the linear regression lines
Contrasts in growth response to drought between age groups
The radial growth of the older trees was more likely to be negatively affected by drought during the growing season. There was a significant (P < 0.05) positive correlation between RWI of the trees in PS2 and June scPDSI over 2010 − 2019 as well as first-order differences over 2011 − 2019 (Fig. 4a). In addition, although no significant correlation occurred between the RWIs of PS2 and scPDSI during the declining period of 2004 − 2019 (correlation results not shown), the first-order differences of the two variables were significantly (P < 0.01) positively correlated over 2005 − 2019 (Fig. 4b), indicating their high frequency associations. For PS1, there was no significant relationship between RRW and monthly scPDSI from January to October during 2010–2019 (Fig. 4a). To further determine the growth-drought relationship over the early period of tree development, a correlation analysis was carried out between RWI and scPDSI during a much longer period 1983–2012 for PS2, a period when the age of most trees was < 30 years (Fig. 1b); with age < 30 years, drought-related growth decline has been rarely observed. There was no significant correlation between these two variables (Fig. S2 in Online Appendix). In addition, there was a significant (P < 0.05) positive correlation between the first-order differences of BAI for PS2 and of June scPDSI (Fig. 4d), but it did not occur for PS1 over the same period (result not shown).

Relationships between the tree-ring metrics and basal area increment (BAI) of the two age groups of P. sylvestris var. mongolica and self-calibrating Palmer drought severity index (scPDSI) as well as between their first-order differences. a correlations with monthly scPDSI from January to October for the raw tree-ring width (RRW) of younger (PS1) and the tree-ring width indices (RWIs) of older (PS2) groups of P. sylvestris var. mongolica during their common period of 2010 − 2019; b variations in the first-order differences of RWIs for PS2 and of June scPDSI during the period of 2005 − 2019 (corresponding to the significant decreasing period of 2004–2019 for RWIs in Fig. 1b); c variations in the first-order differences of RWIs for PS2 and of June scPDSI during the period of 1984 − 2019; d variations in the first-order differences of BAI for PS2 and of June scPDSI during the period of 2009–2019 (corresponding to the common period of 2008–2019 of available BAI data for PS1 and PS2 as shown in Fig. 1c). r1dif denotes the correlation coefficient between the first-order differences of the two variables analyzed. ** denotes that the correlation is statistically significant at the level of 0.05
Stronger responses to drought by radial growth at the individual tree level in older trees relative to younger ones were echoed by the response patterns of their vegetation greenness and water content at the stand level. Both NDVI and NDWI showed higher correlations with July scPDSI during the growing season in PS2 than PS1, although the correlations between the former two variables and July scPDSI were significant (P < 0.05) in both age groups (Fig. 5a, b). In addition, correlations between the first-order differences of NDVI and those of July scPDSI were significant (P < 0.05) in PS2 but not in PS1 (Fig. 5c). There were no differences in correlations for the first-order differences of NDWI and those of July scPDSI between the two groups (Fig. 5d).

Scatter plots showing the relationships of both normalized difference vegetation index (NDVI) and normalized difference water index (NDWI) with July self-calibrating Palmer drought severity index (scPDSI) over their common period of 2016 − 2020 a, b as well as those of their first-order differences during the 2017 − 2020 period (c, d), for the younger (PS1) and older (PS2) groups of P. sylvestris var. mongolica. The solid and dashed lines presented in the sub-panels denote that the correlations between NDVI/NDWI and July scPDSI for the two age groups are statistically significant and insignificant at the level of 0.05, respectively
Differences in xylem hydraulics between young and older age groups
Xylem hydraulic characteristics varied substantially between the two age groups. Both Ks and Kl of the trees in PS2 were significantly (P < 0.1) lower than those in PS1, with a decrease of 49.0 and 40.7%, respectively (Fig. 6a, b). Compared with the younger trees, LA/SA declined in the older trees (Fig. 6c).

Sapwood-area (Ks, a) and leaf-area (Kl, b) specific hydraulic conductivities and leaf area to sapwood area ration (LA/SA, c) respectively for the younger (PS1) and older (PS2) groups of P. sylvestris var. mongolica. Error bar above each column denotes 1 standard error (n = 4). Different and same lowercase letters in each sub-panel denote statistical significance and insignificance at the level of 0.1 for the two age groups, respectively
Discussion
Greater risks of growth declines in aging Mongolian pine under drought
Older, mature trees are more vulnerable to growth decline or to mortality than younger ones, which might be attributed to greater impacts of drought (Lindenmayer et al. 2012; Bennett et al. 2015). Our results show that older Mongolian pines experienced an obvious decline in radial growth and concurrently showed a greater sensitivity to drought during the growing season, as reflected by higher correlations of RWIs and BAI with June scPDSI. This concurs with a tree-ring investigation that found more obvious decrease of radial growth in older trees than younger ones of Qinghai spruce in Qilian Mountains in northwestern China under the influence of climate change (Wang et al. 2021). In addition, using plot-based measurements, several studies have also indicated that aboveground biomass of old forests in western Canada were more strongly affected by climate change relative to younger forests and were particularly vulnerable to growth decline caused by increasing drought associated with climate warming (Chen and Luo 2015; Chen et al. 2016). A synthesized analysis of diameter growth data revealed that, on a global scale, drought played a more detrimental role in radial growth and mortality of larger old-growth trees than smaller ones (Bennett et al. 2015). As large trees play a key role in forest ecosystems and radial growth is closely associated with biomass accumulation (Lamsal et al. 2012; Lindenmayer et al. 2012; Bennett et al. 2015), greater vulnerability in the radial growth to drought indicates that the dynamics of forest carbon sequestration would be largely affected by climate extremes and water availability changes related to climate fluctuations. The larger old-growth trees in Mongolian pine plantations tend to suffer from drought-driven radial growth decline, especially in water-limited environments as in our study area, which could substantially influence the capacity of carbon fixation and other benefits they provide.
Consistent with increasing vulnerability of radial growth with age to drought, canopy greenness and water content at the stand level for older trees were more likely to be negatively affected by drought than younger ones. This is shown by stronger correlations of NDVI and NDWI with the growing season scPDSI in the older plantation. This agrees with a recent finding that old-growth forests at the southern edge of the taiga have low resilience at the canopy level in coping with water stress (Liu et al. 2021). Studies have shown a significant positive correlation between radial growth (i.e., RWIs) and satellite-derived vegetation indices (i.e., NDVI) under similar climatic conditions, especially for the relationships with vegetation indices on seasonal and longer time scales (Leavitt et al. 2008; Bunn et al. 2013; Vicente-Serrano et al. 2016). As there are associations between the variables of the two aspects, they may exhibit synchronized changes in sensitivity to drought with stand age.
There is, however, no close linkage between individual tree radial growth and vegetation greenness and water content at the stand level in relatively short time scales for the plantations analyzed. This is indicated by the lack of correlation of RWIs and BAI with the daily NDVI and NDWI for the same age group. This is consistent with the findings of studies showing no relationship between radial growth and satellite-based measurements at biweekly or daily scales (Beck and Goetz 2011; Berner et al. 2013; Brehaut and Danby 2018). That there were no statistically significant correlations between the metrics of these two aspects might be due to the long-term dynamics of carbon stored in woody parts of trees. This is reflected by radial growth and does not fully depend on recently synthesized carbohydrates from photosynthesis and is hence possibly decoupled from the short-term carbon gain of leaves at the canopy level (Huston and Wolverton 2009; Vicente-Serrano et al. 2016). Consequently, different responses to climate could occur between radial growth and the condition of the canopy vegetation. The annually-resolved RWIs and BAI were particularly sensitive to June scPDSI, while daily NDVI and NDWI during late July to late August were vulnerable to July scPDSI. However, this study also indicated that, in water-limited environments of our study area, both short-term canopy vegetation dynamics at the stand level (i.e., daily NDVI and NDWI) and relatively long-term radial growth at a tree scale (i.e., RWIs and BAI) were more susceptible to drought during the growing season in older Mongolian pines. This result is in accordance with recent reports that growth at both individual tree and stand levels consistently showed declines when the climate became drier in the Sierra Nevada (southwest USA), a water-deficit area (Wong et al. 2021). In regions with water shortages, trees are particularly vulnerable to drought and the negative impacts were probably expressed in growths at different scales (Beck et al. 2011; Subedi et al. 2021). Unlike several studies that relied on a single method (e.g., Carrer and Urbinati 2004; Liu et al. 2018, 2021), this study examined tree performance by combining tree-ring analysis and remotely sensed VIs that provided a more comprehensive understanding on response of tree growth to water stress with increasing age.
Water-related mechanisms on growth decline of aging Mongolian pine
Greater vulnerability of larger old-growth trees to drought-induced growth decline relative to younger trees may be attributed to greater restrictions on water transport. Trees become more susceptible to drought and suffer more severe hydraulic limitations with height increase, as deduced by Darcy’s law of permeability (McDowell and Allen 2015). Compared with smaller, younger trees, larger ones must overcome greater resistance for lifting water from roots to crowns because of increased length in water transport pathways, resulting in more challenging xylem hydraulics (Ryan and Yoder 1997; Ryan et al. 2006; Bennett et al. 2015). Furthermore, old trees having larger crowns and leaf areas relative to younger trees endure greater water loss from elevated transpiration that exacerbates soil moisture deficits and reduce water availability (Duan et al. 2021). As water stress and hydraulic limitations become stronger with age, trees become more sensitive to hydroclimatic conditions leading to more prominent climate signals expressed in measurements of radial growth at individual tree level (i.e., RWIs and BAI) and of vegetation canopy condition at stand level (i.e., NDVI and NDWI) for older individuals (Travis et al. 1990; Carrer and Urbinati 2004).
Larger old-growth Mongolian pines in water-deficit environments susceptible to severe xylem hydraulic limitations under increasing water stress. Decreases in the groundwater table in the study area because of the dramatic expansion of water-consuming agricultural land since the 1950s most likely have led to increased water stress in plants (Zheng et al. 2012). The larger, older Mongolian pines might have more potential to avoid water stress than the smaller, younger trees because their larger root systems allow them access to deeper soil water. However, larger old-growth trees having greater root length and mass are commonly accompanied by increase leaf transpiration (Poorter et al. 2012), which exacerbates water loss through elevated transpiration. These causes make it is less likely to convert the merits in extensive root systems into hydraulic advantages for larger old-growth trees under water deficits (Bennett et al. 2015). Conversely, larger, older trees might suffer more from increasingly severe water stress compared to smaller, younger ones as the former depend more on groundwater that has continuously decreased over recent decades (Zheng et al. 2012; Song et al. 2015, 2016). Furthermore, larger old-growth trees may also experience greater hydraulic dysfunction due to the effects of height-related water transport resistance (Ryan and Yoder 1997; Midgley 2003). Such deduction is confirmed by our results that lower Ks and Kl occurred in larger old-growth Mongolian pines relative to the smaller, younger ones. This agrees with Liu et al. (2018) who found increasing hydraulic constraints with tree size for Mongolian pine trees in the vicinity of our study area. Because of greater hydraulic constraints, larger and older Mongolian pines have probably undergone more severe growth decline at both individual tree and stand levels, as indicated by the results of our tree-ring metrics and remotely sensed vegetation indices investigation. Under a future drier climate, we speculate that aging Mongolian pine plantations would be more vulnerable to drought-driven forest die-back.
Conclusions
By integrating tree-ring analysis and remotely sensed vegetation indices, our study suggests that recent decreases in RWIs (radial width indices) at individual tree level was echoed by consistently lower multi-year values of satellite-derived normalized difference vegetation and water indices at the stand level in plantations. In addition, all growth-related metrics were more sensitive to drought stress during the growing season, reflected by strong correlations between these metrics and the June/July scPDSI as well as their first-order differences, in older stands. Consistently lower sapwood-specific and leaf-specific hydraulics occurred in the older trees, indicating that changes of xylem hydraulics played an important role in mediating growth performances to drought. By integrating multiple approaches for quantifying tree growth performance, our study provides a comprehensive understanding of the differences in growth between older and younger Mongolian pine plantations and reveals the underlying causes from xylem hydraulics. Under climate warming, old-growth Mongolian pine plantations will exhibit more pronounced growth decline associated with increased water stress. Therefore, renewing old-growth plantations and taking action to alleviate competitions for water through thinning might be required for the sustainable management of Mongolian pine plantations.
References
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim JH, Allard G, Running SW, Semerci A, Cobb N (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecol Manag 259:660–684
Barichivich J, Osborn TJ, Harris I, van der Schrier G, Jones PD (2020) Drought in “State of the Climate in 2019.” B Am Meteorol Soc 101:1–429
Beck PSA, Goetz SJ (2011) Satellite observations of high northern latitude vegetation productivity changes between 1982 and 2008: ecological variability and regional differences. Environ Res Lett 6:045501
Beck PSA, Juday GP, Alix C (2011) Changes in forest productivity across Alaska consistent with biome shift. Ecol Lett 14:373–379
Bennett AC, McDowell NG, Allen CD, Anderson-Teixeira KJ (2015) Larger trees suffer most during drought in forests worldwide. Nat Plants 1:15139
Berner LT, Beck PSA, Bunn AG, Goetz SJ (2013) Plant response to climate change along the forest-tundra ecotone in northeastern Siberia. Global Change Biol 19:3449–3462
Biondi F, Qeadan F (2008) A theory-driven approach to tree-ring standardization: defining the biological trend from expected basal area increment. Tree-Ring Res 64:81–96
Brehaut L, Danby RK (2018) Inconsistent relationships between annual tree ring-widths and satellite measured NDVI in a mountainous subarctic environment. Ecol Indic 91:698–711
Bunn AG, Hughes MK, Kirdyanov AV, Losleben M, Shishov VV, Berner LT, Oltchev A, Vaganov EA (2013) Comparing forest measurements from tree rings and a space-based index of vegetation activity in Siberia. Environ Res Lett 8:035034
Carrer M, Urbinati C (2004) Age-dependent tree-ring growth responses to climate in Larix decidua and Pinus cembra. Ecology 85:730–740
Chen HY, Luo Y (2015) Net aboveground biomass declines of four major forest types with forest ageing and climate change in western Canada’s boreal forests. Global Change Biol 21:3675–3684
Chen DY, Huang JF, Jackson TJ (2005) Vegetation water content estimation for corn and soybeans using spectral indices derived from MODIS near- and short-wave infrared bands. Remote Sens Environ 89:225–236
Chen HYH, Luo Y, Reich PB, Searle EB, Biswas SR (2016) Climate change-associated trends in net biomass change are age dependent in western boreal forests of Canada. Ecol Lett 19:1150–1158
Choat B, Brodribb TJ, Brodersen CR, Duursma RA, Lopez R, Medlyn BE (2018) Triggers of tree mortality under drought. Nature 558:531–539
Ciais P, Reichstein M, Viovy N (2005) Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 437:529–533
Cook ER, Kairiukstis LA (1990) Methods of dendrochronology. Kluwer Academic Press, Dordrecht, Netherlands
Cook ER (1985) A time-series analysis approach to tree-ring standardization. Tucson Ph.D. Thesis, University of Arizona Press
Drusch M, Del Bello U, Carlier S (2012) Sentinel-2: ESA’s optical high-resolution mission for GMES operational services. Remote Sens Environ 120:25–36
Duan CY, Li MY, Fang LD, Cao Y, Wu DD, Liu H, Ye Q, Hao GY (2021) Greater hydraulic safety contributes to higher growth resilience to drought across seven pine species in a semi-arid environment. Tree Physiol 42:727–739
Enquist BJ (2003) Cope’s rule and the evolution of long-distance transport in vascular plants: allometric scaling, biomass partitioning and optimization. Plant Cell Environ 26:151–161
ERDAS (1999) ERDAS field guide. Atlanta, Georgia
Feng JC, Jiang J (1996) On the historical changes of horqin sandy land. J Chin Histor Geogr 4:105–120 (in Chinese)
Fritts HC (1976) Tree-rings and climate. Academic Press, London
Gao B (1996) A normalised difference water index for remote sensing of vegetation liquid water from space. Remote Sens Environ 58:257–266
Gascon F, Bouzinac C, Thépaut O, Jung M, Francesconi B, Louis J, Lonjou V, Lafrance B, Massera S, Gaudel-Vacaresse A, Languille F, Alhammoud B, Viallefont F, Pflflug B, Bieniarz J, Clerc S, Pessiot L, Trémas T, Cadau E, De Bonis R, Isola C, Martimort P, Fernandez V (2017) Copernicus sentinel-2A calibration and products validation status. Remote Sens 9:584
Gora EM, Esquivel-Muelbert A (2021) Implications of size-dependent tree mortality for tropical forest carbon dynamics. Nat Plants 7:384–391
Hogg EH, Michaelian M, Hook TI, Undershultz ME (2017) Recent climatic drying leads to age-independent growth reductions of white spruce stands in western Canada. Global Change Biol 23:5297–5308
Huston MA, Wolverton S (2009) The global distribution of net primary production: resolving the paradox. Ecol Monogr 79:343–377
SPSS Inc (1997) SPSS Advanced statistics: statistical package for the social sciences. Adv Statist 7.5. Chicago, IL: SPSS Inc
IPCC (2013) Climate Change 2013: The Physical Science Basis (eds Stocker TF et al.). Cambridge Univ Press, Cambridge
IPCC (2021) AR6 Climate Change 2021: The physical science basis. The working group I contribution to the sixth assessment report of the intergovernmental panel of climate change, Geneva, Switzerland
Ji L, Zhang L, Wylie B, Rover J (2011) On the terminology of the spectral vegetation index (NIR − SWIR) / (NIR + SWIR). Int J Remote Sens 32:6901–6909
Kannenberg SA, Maxwell JT, Pederson N, D’Orangeville L, Ficklin DL, Phillips RP (2019) Drought legacies are dependent on water table depth, wood anatomy and drought timing across the eastern US. Ecol Lett 22:119–127
Karnieli A, Agam N, Pinker RT, Anderson M, Imhoff ML, Gutman GG, Panov N, Goldberg A (2010) Use of NDVI and land surface temperature for drought assessment: merits and limitations. J Clim 23:618–633
Kogan FN (1990) Remote sensing of weather impacts on vegetation in non-homogeneous areas. Int J Remote Sens 11:1405–1419
Korhonen L, Packalen P, Rautiainen M (2017) Comparison of Sentinel-2 and Landsat 8 in the estimation of boreal forest canopy cover and leaf area index. Remote Sens Environ 19:259–274
Lamsal S, Rizzo DM, Meentemeyer RK (2012) Spatial variation and prediction of forest biomass in a heterogeneous landscape. J Forestry Res 23:13–22
Leavitt SW, Chase TN, Rajagopalan B, Lee E, Lawrence PJ (2008) Southwestern U.S. tree-ring carbon isotope indices as a possible proxy for reconstruction of greenness of vegetation. Geophys Res Lett 35:L12704
Lindenmayer DB, Laurance WF, Franklin JF (2012) Global decline in large old trees. Science 338:1305–1306
Liu YY, Wang AY, An YN, Lian PY, Wu DD, Zhu JJ, Meinzer FC, Hao GY (2018) Hydraulics play an important role in causing low growth rate and dieback of aging Pinus sylvestris var. mongolica trees in plantations of Northeast China. Plant Cell Environ 41:1500–1511
Liu F, Liu HY, Xu CY, Shi L, Zhu XR, Qi Y, He WQ (2021) Old-growth forests show low canopy resilience to droughts at the southern edge of the taiga. Global Change Biol 27:2392–2402
Luo Y, Chen HYH (2013) Observations from old forests underestimate climate change effects on tree mortality. Nat Commun 4:1655
McDowell NG, Allen CD (2015) Darcy’s law predicts widespread forest mortality under climate warming. Nat Clim Change 5:669–672
McMahon SM, Parker GG, Miller DR (2010) Evidence for a recent increase in forest growth. Proc Natl Acad Sci USA 107:3611–3615
Midgley JJ (2003) Is bigger better in plants? The hydraulic costs of increasing size in trees. Trends Ecol Evol 18:5–6
Petit G, Anfodillo T, Carraro V, Grani F (2011) Hydraulic constraints limit height growth in trees at high altitude. New Phytol 189:241–252
Poorter H, Niklas KJ, Reich PB, Oleksyn J, Poot P, Mommer L (2012) Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control: Tansley review. New Phytol 193:30–50
Rozas V, DeSoto L, Olano JM (2009) Sex-specific, age-dependent sensitivity of tree-ring growth to climate in the dioecious tree Juniperus thurifera. New Phytol 182:687–697
Ryan MG (2015) Tree mortality: large trees losing out to drought. Nat Plants 1:15150
Ryan MG, Yoder BJ (1997) Hydraulic limits to tree height and tree growth. Bioscience 47:235–242
Ryan MG, Phillips N, Bond BJ (2006) The hydraulic limitation hypothesis revisited. Plant Cell Environ 29:367–381
Schweingruber FH (1988) Tree ring—basic and applications of dendrochronology. Paul Haupt Press, Berne
Sippel S, Reichstein M, Ma XL, Mahecha MD, Lange H, Flach M, Frank D (2018) Drought, heat, and the carbon cycle. Curr Clim Change Rep 4:266–286
Song LN, Zhu JJ, Yan QL, Li MC, Yu GQ (2015) Comparison of intrinsic water use efficiency between different aged Pinus sylvestris var. mongolica wide windbreaks in semiarid sandy land of northern China. Agroforest Syst 89:477–489
Song LN, Zhu JJ, Li MC, Zhang JX (2016) Water use patterns of Pinus sylvestris var. mongolica trees of different ages in a semiarid sandy lands of Northeast China. Environ Exp Bot 129:94–107
Sperry JS, Tyree MT (1988) Mechanism of water stress-induced xylem embolism. Plant Physiol 88:581–587
Subedi MR, Xi WM, Edgar CB, Rideout-Hanzak S, Yan M (2021) Tree mortality and biomass loss in drought-affected forests of East Texas, USA. J Forestry Res 32:67–80
Tao Y (1987) Historico-ecological development of Keerqin sandy land and lessons from it. Chin J Ecol 6:39–42 (in Chinese)
Travis DJ, Meentemeyer V, Belanger RP (1990) Stressed trees produce a better climatic signal than healthy trees. Tree-Ring Bulletin 50:29–32
Tucker CJ (1979) Red and photographic infrared combinations for monitoring vegetation. Remote Sens Environ 8:127–150
Tyree MT (2003) Hydraulic limits on tree performance: transpiration, carbon gain and growth of trees. Trees-Struct Funct 17:95–100
Tyree MT, Ewers FW (1991) The hydraulic architecture of trees and other woody plants. New Phytol 119:345–360
Ukkola AM, De Kauwe MG, Roderick ML, Burrell A, Lehmann P, Pitman AJ (2021) Annual precipitation explains variability in dryland vegetation greenness globally but not locally. Global Change Biol 27:4367–4380
van der Schrier G, Barichivich J, Briffa KR, Jones PD (2013) A scPDSI-based global data set of dry and wet spells for 1901–2009. J Geophys Res Atmos 118:4025–4048
Vicente-Serrano SM, Gouveia C, Camarero JJ, Beguería S, Trigo R, López-Moreno JI, Azorín-Molina C, Pasho E, Lorenzo-Lacruz J, Revuelto J (2013) Response of vegetation to drought time-scales across global land biomes. Proc Natl Acad Sci 110:52–57
Vicente-Serrano SM, Camarero JJ, Olano JM, Martín-Hernández N, Peña-Gallardo M, Tomás-Burguera M, Gazol A, Azorin-Molina C, Bhuyan U, Kenawy AE (2016) Diverse relationships between forest growth and the normalized difference vegetation index at a global scale. Remote Sens Environ 187:14–29
Wang B, Yu PT, Yu YP, Zhang L, Wang YH, Wan YF, Yang WJ, Wang SL, Liu XD (2021) Response of radial growth of Qinghai spruce at different ages to climate change in Qilian Mountains, Northwestern China. Scientia Silvae Sinicae 57:1–8
Wells N, Goddard S, Hayes MJ (2004) A self-calibrating palmer drought severity index. J Clim 17:2335–2351
West H, Quinn N, Horswell M (2019) Remote sensing for drought monitoring and impact assessment: progress, past challenges, and future opportunities. Remote Sens Environ 232:111291
Whitehead D, Jarvis PG (1981) Water Deficits and Plant Growth (ed. Kozlowski TT). Academic Press, London
Wigley T, Briffa KR, Jones PD (1984) On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J Clim Appl Meteorol 23:201–213
Wong CYS, Young DJN, Latimer AM, Buckley TN, Magney TS (2021) Importance of the legacy effect for assessing spatiotemporal correspondence between interannual tree-ring width and remote sensing products in the Sierra Nevada. Remote Sens Environ 265:112635
Zheng X, Zhu JJ, Yan QL, Song LN (2012) Effects of land use changes on the groundwater table and the decline of Pinus sylvestris var. mongolica plantations in southern Horqin Sandy Land. Northeast China Agr Water Manage 109:94–106
Zhu JJ, Kang HZ, Tan H, Xu ML, Wang J (2005a) Natural regeneration characteristics of Pinus sylvestris var. Mongolica forests on sandy land in Honghuaerji. China J Forestry Res 16:253–259
Zhu JJ, Kang HZ, Tan H, Xu ML (2006) Effects of drought stresses induced by polyethylene glycol on germination of Pinus sylvestris var. mongolica seeds from natural and plantation forests on sandy land. J Forestry Res 11:319–328
Zhu JJ, Zheng X, Yan QL (2016) Assessment of impacts of the three-north protective forest program on ecological environments by remote sensing technology–Launched after 30 years (1978–2008). Science Press, Beijing (in Chinese)
Zhu JJ, Zeng DH, Kang HZ, Wu XY, Fan ZP (2005b) Decline of Pinus sylvestris var. mongolica lantations on Sandy Land. China Forestry Publishing House, Beijing (in Chinese).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding: This study was financially supported by the National Natural Science Foundation of China (31901093, 32220103010, 32192431, 31722013), National Key R & D Program of China (2020YFA0608100, 2022YFF1302505) and the Key Research Program of Frontier Sciences of the Chinese Academy of Sciences (ZDBS-LY-DQC019).
The online version is available at http://www.springerlink.com.
Corresponding editor: Tao Xu
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Li, M., Yang, L., Cao, Y. et al. Aging Mongolian pine plantations face high risks of drought-induced growth decline: evidence from both individual tree and forest stand measurements. J. For. Res. 35, 38 (2024). https://doi.org/10.1007/s11676-023-01677-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11676-023-01677-4