
Abstract
When considering deficit irrigation, the most resistant period to water stress in olive crops is during the pit hardening. However, the determination of this period and the impact that water stress has on its onset or duration have not been clearly established. The aim of this work is to describe the impact of water stress on three processes that determine fruit development: fruit growth, endocarp hardening, and oil accumulation. This experiment was conducted between 2021 and 2022, in a plot with olive trees that were 3 years old at the beginning of the experiment. The young trees were of cv Manzanilla de Sevilla and were planted in a 4 × 1.5 m frame. The treatments were: Control, irrigated close to what is considered the optimum level in which the water status of the crop ensures the absence of water stress; RDI, with a moderate stress during the pit hardening stage; Rainfed and Traditional Rainfed. This latter was included only in 2022 using a 7 × 5 m frame. Traditional Rainfed was included to compare with Rainfed, these were similar in the maximum level of water stress but not in the duration. The fruit growth, oil accumulation, and endocarp hardness were measured for a period of 2 years. The hardness curves fit accurately established Period II which was the one with the fast hardness increases. The water potential was measured periodically on leaves at midday, whereby the minimum and average potential and the stress integral were obtained for each of the periods indicated before using pit hardness measurements. The results suggest that endocarp hardening was the process most resistant to water stress, as it showed very little variation from the seasonal pattern, despite the high levels of stress achieved. The fruit growth process was the most sensitive to water stress, but the recovery was fast in conditions of moderate water stress level. The oil accumulation started slowly during Period II, but its rate increased at the end of the period. This later process was moderately resistant to water stress. However, although the accumulation rate was faster during Period III, it was influenced by the water status during Period II. Cumulative water stress was related with fruit growth and oil accumulation processes. Endocarp development could be useful for determining a/the deficit irrigation period because of its almost constant pattern. These periods would permit water management optimization according to final use of the fruit (oil or table). However, continuous monitoring of water stress would be needed.
Similar content being viewed by others


Introduction
Water is becoming an increasingly limited resource due to reduced meteorological precipitation and rising temperatures, which results in greater demand and a surge in non-agricultural uses (Lorite et al. 2022). For these reasons, farmers are forced to use deficit irrigation scheduling. The aim of regulated deficit irrigation (RDI) is to impose water stress at times where the impact on the yield is very small or inconsequential (Chalmers and Wilson 1978). Irrigation scheduling may cause a yield reduction when implemented without taking into account the phenological stage (e.g., peach, Girona et al. 2012) or the water stress level imposed (e.g., olive, Goldhamer 1999).
Olive (Olea europaea L.) trees have a high resistance to water stress (Díaz-Espejo et al. 2018), and their endocarp hardening period is considered the time where such stress has the lowest impact on the final yield (Goldhamer 1999). Nevertheless, the final result of water stress on the yield depends on the point in time in which it is applied, as well as on its intensity and duration (Hsiao 1990). For olive trees, this is even more complex because the production may be intended for different uses, such as table olives, where the most important parameter is fruit size, or oil production. This involves diverse irrigation strategies, since the sensitivity of the affected processes is also different. The fruit growth process is the most sensitive (Martin-Palomo et al. 2020), and excessively harsh water stress conditions (Goldhamer 1999) or insufficient rehydrations (Corell et al. 2020) may result in a yield loss. Conversely, oil accumulation is a more resistant process, and it has even been reported as being minimally affected by water stress (Navas-Lopez et al. 2019; López-Bernal et al. 2021). However, some studies indicated that there was indeed some sensitivity in oil-producing crops (Fernández et al. 2013; Ahumada-Orellana et al. 2017; Corell et al. 2022).
Therefore, RDI studies with olive trees tend to use endocarp/pit hardening as a reference, although few actually made measurements to estimate this period, considering fixed dates to establish the water stress application instead (e.g., Goldhamer 1999; Moriana et al. 2003). On occasion, this phenological stage has been described as the time when it is not possible to “cut the fruit with a knife” (e.g., Garcia-Inza et al. 2014). But this type of measurement indicates the end of the hardening stage, not its beginning, and this is a serious simplification. Rapoport et al. (2004) described the endocarp hardening period as the gradual appearance of sclereids that start at the time of pollination. Rapoport et al. (2013) used three different periods to describe the evolution of the endocarp hardening. The sclerification process is initially very slow (Period I), but the rate increases once the endocarp reaches its maximum size (Period II), until reaching its maximum hardness (Period III) (Rapoport et al. 2013). Consequently, Period II of fast hardness growth would be the stage with the greatest resistance to water stress.
Recently, Sánchez-Piñero et al. (2022) monitored hardness in different campaigns of different cultivars, in different locations and in the absence of water stress and reached the conclusion that the beginning of Period II takes place 49 days after full bloom. This duration was similar to that suggested by Rapoport et al. (2013) for the Cornicabra cultivar and it could be considered a constant for the species. However, the duration of Period II was variable and dependent on the temperature and the cultivar considered (Sánchez-Piñero et al. 2022). This period was the most common to suggest irrigation restrictions (i.e., Goldhamer 1999). Therefore, if the length changed, water stress could be applied in the most sensitive times of fruit development (such as oil accumulation). In addition, the changes in the pattern of Period II could also be affected by water stress. If the duration of these periods was modified, the real impact of RDI strategies would be uncertain. Hammani et al. (2013) suggested that the beginning of Period II would be moved forward under water stress conditions, while Rapoport et al. (2004) indicate the opposite.
The impact of water stress on yield has multi-year components (shoot growth) as well as others within the same year (fruit size, oil amount). These latter processes would be affected at the time when water stress is applied. Because endocarp development was not commonly described, the real effect of water stress on these yield components is unknown. The aim of this work was to determine the impact of water stress on the different processes taking place in the fruit (endocarp formation, fruit growth, and oil accumulation), and accurately measured the pit hardening changes to establish three development periods. In addition, the water stress impact was assessed considering point thresholds (maximum stress values) or cumulative thresholds (water stress duration and intensity). The description of these responses will permit the identification of possible limitations in the use of pit hardening as a reference in deficit irrigations and the response of this yield component to water stress.
Materials and methods
Site description and experiment design
The experiment was conducted during the years 2021 and 2022 in an experimental plot at the farm “La Hampa”, in Coria del Río (Seville, Spain, 37º 17’N, 6º 3’W, 30 m). The cultivar used was “Manzanilla de Sevilla,” and the trees were 3 years old at the beginning of the experiment, planted at a distance of 4 × 1.5 m. The first harvest took place during the 2020 campaign and the yield was 800 kg ha−1. The soil depth was variable but always deeper than 1 m and had a sandy-loam composition. The irrigation water came from a well located on the farm, and it was pumped to a dam before it entered the irrigation system. The mineral composition of the water was adequate for olive irrigation (pH 8.3; 2.37 dS m−1; 73 mg l−1 Ca; 101 mg l−1 Mg; 225 mg l−1 Na).
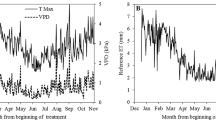
Climate data were collected from the weather station at “La Puebla del Rio” (SIAR 2023). This station belongs to the Andalusian network and is located 6 km away from the site. Figure 1 shows the reference evapotranspiration (ETo) and rainfall during the experiment. The seasonal patterns for both variables are typical for a Mediterranean climate in which rains fall mainly in winter and spring and summer is very dry (Fig. 1).

Seasonal pattern of reference evapotranspiration (ETo) and rain during the two seasons of the experiment. Source: “La Puebla del Rio” station (SIAR 2023). Vertical lines limited the experimental period. Arrows marked the full bloom date
ETo values reached their peak from the end of spring to the beginning of autumn, standing above 5 mm day−1 on some days. During the period of the experiment, the average measured ETo was above this value. The amount of seasonal rainfall during both years was below the average for the area in the last decade (534 mm (AEMET 2023)), and it was approximately 355 mm, 65% of the long-term average. There was an almost complete absence of rainfall during the experiment period in both years, which is frequent in the area. The below-average rainfall meant that the period immediately before flowering was drier than usual, particularly in 2021 (Fig. 1).
The experiment design consisted of randomized plots with three repetitions per treatment. Each experimental plot included 5 rows of 13 trees each, and the 3 central rows were used for the measurements. The irrigation system consisted of a simple line of drips of 2.2 l h−1 at a distance of 0.5 m. Irrigation treatments started in 2020 and were based on the combination of the water stress application time point and intensity. The irrigated season was divided in three different phenological stages in which pit hardening was the main reference. Before and after pit hardening periods were considered as sensitive to water stress in relation to yield respond. The former included the main period of vegetative growth, full bloom, and fruit set. The latter occurred in a few weeks before harvest in this table cultivar green style preparation and included the rehydration period in deficit irrigation management. The start of pit hardening was estimated, according to Rapoport et al. (2013), when the rate of olive longitudinal growth started decreasing. The end of this period, in both years, was a fixed date on the third week of August. The irrigation treatments were:
-
Control. Irrigation scheduling based on the midday stem water potential (SWP) measurements and the estimated evapotranspiration of the crop (ETc). ETc was calculated using the FAO methodology, with a crop coefficient (Kc) of 0.55 and a reduction coefficient (Kr) of 0.7 in 2021 and 1 in 2022. In 2021, the intention was to optimize vegetative growth to achieve maximum yield in the shortest time possible. To that end, the applied water was 100% ETc, except when SWP measurements were more negative than −1.0 MPa, when they were increased to 175%. In 2021, the yield obtained from irrigated treatments was similar to that from an adult plot (approximately 10,000 kg.ha−1), so in 2022, the irrigation scheduling was modified to prevent an excess of vigor. During the second year, the increase in applied water during the pit hardening period was only made when SWP was 0.5 MPa more negative than the baseline established by Corell et al. (2016). This strategy meant that the total amount of water applied was 772 mm in 2021 and 727 mm in 2022.
-
Regulated deficit irrigation (RDI). Irrigation scheduling was also based on SWP measurements, but different thresholds were taken into account in both years according to the phenological stage. In 2021, the SWP threshold values were those estimated using the baseline from Corell et al. (2016), except during pit hardening, when the threshold was fixed at −2.0 MPa for the entire period. In 2022, the SWP threshold was obtained from the baseline by Corell et al. (2016) minus 0.5 MPa, except during the hardening period, when the differential was increased up to 1.5 MPa. The applied water varied based on the difference between the SWP measured and the threshold at any given time. When the SWP was more negative than the threshold, irrigation started with 1 mm day−1, then increased to 2, 3, and 4 mm day−1 when the differential was higher than 10, 20, or 30%, respectively. This scheduling meant that the total amount of water applied was 334 mm in 2021 and 200 mm in 2022.
-
Rainfed. The treatment was not irrigated in June 2020 and received 45 mm of complementary irrigation in September of that year. In 2021 and 2022, no irrigation was provided at all.
-
Traditional Rainfed. In 2022, a Traditional Rainfed plot with the same cultivar in a 7 × 5 m plantation and 24 years of age at the beginning of the trial was assessed. The water stress in this treatment was shorter but similar in water stress level than the previous one. This permitted a comparison in the effect of the duration.
The fruits yield in both years showed a very marked alternate bearing pattern. In 2021, the irrigation treatments produced an average yield close to 10,000 kg ha−1, while in 2022, the yield was between 500 and 1000 kg ha−1 in all treatments, except for Traditional Rainfed, which was approximately 2500 kg ha−1.
Measurement description
The endocarp development was monitored using weekly measurements of pit hardness. Two weeks after full bloom, random samples were taken from ten fruits per repetition of each treatment. The breaking pressure was determined using a device similar to that described in Rapoport et al. (2013). In short, this equipment allows using manual pressure to convert it into the vertical movement of a probe with a 2 mm diameter tip. This tip is connected to a load cell that allows for measuring the force applied. The load cell electrical signal was sent to a data receiver (Model KUSB 318, Keithley Instruments Inc., Cleveland, OH, USA) connected to a computer where data were stored. Data collected during treatment were adjusted to a sigmoid equation:
where PBP is the pit-breaking pressure, a is the range from the minimum to the maximum PBD, b is the slope of the inflection point, c is the date when the inflection point occurs, d is the minimum PBD and DAFB are the days after full bloom.
Adjusting this equation allowed for the definition of the three development periods described by Rapoport et al. (2013): Period I, slow hardness increase; Period II, fast growth; and Period III, maximum hardness values. To determine the beginning of these periods for each of the samples obtained, Eq. 1 was adjusted for each repetition, so the starting and end dates of Period II were established when the equation estimated that 10% and 95% of hardness were reached, respectively.
Determining the amount of oil in the fruit was done by randomly sampling ten olives per repetition on the same date of endocarp development sampling. Fresh and dry weights were determined after reaching constant weight in an oven at 65 ºC. The determination of the oil content was done using a sample from ten dried fruits and a nuclear magnetic resonance spectroscopy (NMR, Minispec, Bruker, Massachusetts, USA). The amount of oil per fruit was determined by multiplying the dry weight by the percentage of oil obtained.
The olive size was determined weekly using ten fruits per treatment, selected randomly. These fruits were measured while still on the trees by taking the length and width in the middle area to calculate the volume.
The water status of the trees was estimated through a SWP measurement per repetition. The SWP was measured on fully expanded leaves that were covered at least 2 h before taking the measurement using a pressure chamber (Model 1000, PMS, USA). SWP data were analyzed to obtain three indicators in different phenological stages: minimum SWP, average SWP, and stress integral (SI). The SI was estimated using the modified Myers formula (1988), where the maximum value was different on each date using the one estimated with the baseline from Corell et al. (2016):
where SI is the stress integral, SWP is the midday stem water potential, BL is the value estimated on the baseline, and n is the number of days.
The periods where these indicators were calculated included pre-flowering, Period I, II, and III of endocarp development, according to the estimations based on the hardness measurements.
Statistical analysis
The statistical analysis was conducted with ANOVA and a separation of means (Tukey test) using the Statistic (SX, v 8.0) program. Significant differences with a p level <0.05 were considered in both tests. The assessment of the water stress effect on some parameters was done by obtaining the linear regression between them and the indicators calculated for each period. Multivariate analyses were conducted, and different indicators and/or periods were included; however, they did not improve simple fits. Sigmoid parameters were estimated with nonlinear regression tools with the package “nls2” in R environment (R Core team 2013).
Results
Endocarp development and water status
Pit hardening followed a sigmoid pattern in all treatments, both in the 2021 season (Fig. 2a) and in the 2022 season (Fig. 2b), due to a variation of the pit hardening rate, which remained constant during an initial period (Period I), then started a fast increase (Period II), and ended when the maximum value was reached (Period III).

Seasonal pattern of pit-breaking pressure curve during the 2021 (a) and 2022 (b) seasons. Blue circle: Control; green triangle: RDI; red square: Rainfed; yellow diamond: Traditional Rainfed (colour figure online)
Afterward, it remained constant for the rest of the fruit’s growth and ripening. The pattern change was the same for all treatments, except the Rainfed one in 2022 (Fig. 2b). This treatment showed maximum seasonal values that were almost constant from day of year (DOY) 190 and clearly below the others. Table 1 shows the parameters for the individual fits of the sigmoid equation for these pit hardening curves.
In 2021, there were no significant differences in any of these parameters for the different treatments. The number of days until half the maximum hardness is achieved (Table 1, parameter c) was very similar, approximately 71 days from full bloom. The hardening rate during Period II (Table 1, parameter b) was also similar at approximately 0.13 MPa day−1, with maximum hardness (Table 1, a + d) at around 165 MPa. Conversely, the Rainfed treatment displayed a maximum hardness, which was significantly lower (108.4 MPa) than the rest in 2022. Even if the rest of the parameters did not show any important variations, the pit hardening rate (b) tended to have higher values than the rest, and the date when 50% of the hardening value was reached (c) tended to have lower values in the Rainfed treatment. The RDI treatment in 2022 also tended to have values suggesting a faster hardening curve (steeper slope (b), fewer days (c)) and lower hardness (a + d) than the rest, although the differences were much smaller than those seen in the Rainfed treatment (Table 1).
The curve fits in Fig. 2 and Table 1, together with the bloom date, allowed for estimating the duration of endocarp development Periods I and II (Table 2).
In 2021, full bloom occurred on DOY 107 in all treatments, whereas in 2022, this took place on DOY 111 in the Rainfed and Traditional Rainfed treatments, and on DOY 118 in Control and RDI treatments. The duration of Periods I and II in the 2021 season was similar for all treatments, and the differences were not significant, as they were never longer than 2 days. Conversely, the duration of Period I in 2022 for the Rainfed treatment was considerably shorter than the rest, and the difference was more than 10 days. However, in the Traditional Rainfed treatment, this Period was of a similar duration than in RDI and Control and ended significantly earlier than Rainfed. Even though there were no differences in the duration of Period II this year, RDI tended to have lower values than the rest, with 6–8 days of difference vs. Control and the two Rainfed treatments.
The data in Fig. 2 and Table 2 allowed for the establishment of four different periods according to the endocarp development. The water stress indicators for these periods changed throughout the season (Table 3). Measurements during the 2021 pre-flowering period were scarce, and only one was completed due to rainfall (Fig. 1). During Period I, meaningful differences were found between Rainfed and Control only in the SI, although the most negative values for the other indicators were also found in this treatment. On the other hand, the minimum potential (Min-SWP) in Period II was considerably lower in the Rainfed treatment than in the rest. Despite the lack of important differences in the rest of the indicators during this period, the values suggest a very severe stress level in Rainfed vs. Control. For both periods, RDI was an intermediate treatment, without major differences, although with a clear trend toward higher water stress values than Control. Conversely, all indicators showed important differences during Period III. The Rainfed treatment was markedly more stressed than the other two, with values suggesting very severe water stress. Nevertheless, only the SI values could be used to differentiate the three treatment levels during this period. The RDI treatment values were similar during Periods II and III, with the differential increasing in all indicators vs. Control during Period III. The way RDI irrigation scheduling was managed, taking into account a fixed date for rehydration, resulted in the most severe stress conditions for this treatment occurring at the beginning of Period III.
The data for water stress indicators in 2022, on the other hand, showed meaningful differences during all endocarp development periods, but not during pre-flowering (Table 3). During pre-flowering, both RDI and Rainfed showed a trend toward worse indicators, particularly the SI, that was almost double, and in Min-SWP, that clearly dropped in Rainfed. Traditional Rainfed was similar to Control due to the lower density of trees. Period I showed important differences in the SI and Av-SWP, with the RDI treatment being more stressed than Control. Values for both Rainfed treatments were statistically in the middle, although they were closer to RDI and showed values twofold those of Control. This trend was also noticed in Min-SWP, but with no significant differences. The stress level during this Period I was higher than that obtained during the same period in the previous year. Period II showed considerable differences in all indicators, and there were two clearly defined groups: Control and RDI with less stressed values, and Rainfed and Traditional Rainfed, with severe levels. Although there were no significant differences between RDI and Control, the latter tended to have lower stress values than the former. During Period II, indicators for the two Rainfed treatments suggested higher water stress levels than those obtained the previous year, as opposed to the irrigation treatments. Lastly, the differences in Period III changed according to the indicator considered. Only Av-SWP displayed differences for the four treatments, while the rest showed statistical differences in the treatments with the most different values. In all cases, there were meaningful differences between the Control and the Rainfed treatments. The RDI and the Traditional Rainfed treatments were statistically in the middle. The RDI treatment was closer to Control, but with very negative Min-SWP values (−4.35 MPa), while Traditional Rainfed was closer to Rainfed. In this Period III, the SI values were slightly lower than those for 2021, while SWP values were more negative than the previous year, suggesting more severe individual stress values that did not last as long.
Water stress impact on fruit volume growth
The seasonal pattern of fruit volume was similar in the two experimental years (Fig. 3). The seasonal pattern could be likened to a sigmoid, like that of endocarp development, as there are two slowing-down periods, at the beginning and at the end. The latter was clearly noticeable in both Rainfed treatments, but not so much in the irrigation treatments due to the earlier harvest. Nevertheless, there was a gentler upward slope in the last 4 data points of the Control treatment in the discharge year (2022, Fig. 3b), suggesting the beginning of this period. In both seasons, Rainfed and Traditional Rainfed presented clear differences with Control and RDI. For the RDI treatment, the growth slope was gentler than Control from the middle of Period II in both years. However, there were only meaningful differences in fruit growth between Control and RDI in August 2022 (Fig. 3b), although there was a clear trend toward size reduction from the middle of Period II in both seasons. At the beginning of Period III in both years, the RDI treatment data showed a volume drop and a later stop, which resulted in the greatest differences in 2021 (Fig. 3a) and important differences in 2022 (Fig. 3b). This drop was followed by a rehydration that caused the final RDI values to be very similar to those of Control. Conversely, in the Rainfed and Traditional Rainfed treatments, the fruit growth stopped much earlier than RDI, on DOY 176 in 2021 (Fig. 3a) and between DOY 160 and DOY 167 in 2022 (Fig. 3b), and they did not show a fruit volume loss as clearly as in the RDI treatment.

Seasonal pattern of fruit volume during 2021 (a) and 2022 (b) seasons. Each point is the average of three data. Vertical bars represent the standard error. Control, blue circle; RDI, green triangle; Rainfed, red square; Traditional Rainfed, yellow diamond (only 2022). Asterisks indicate significant differences in the dates (Turkey test; p < 0.05). Vertical dashed lines limit the period of massive pit hardening (Period II) as average of all treatments. b Adapted from de Sosa et al. (2023) (colour figure online)
Oil accumulation
The trend of oil accumulation was similar to a sigmoid, although, due to the harvest date for table olives, it did not reach maximum values in the irrigated treatments (Fig. 4). The increase in fruit oil content started to be significant in all treatments toward the middle of Period II, massive pit hardening, in the 2 years considered (Fig. 4). After this process started, the increase in oil content increase was continuous, but the rate was different for the different treatments. In 2021, there were important differences in the fruit oil content between Rainfed and the other two treatments from the end of Period II until the end of the experiment (Fig. 4a). However, there were no meaningful differences between the Control and RDI treatments. During 2022 (Fig. 4b), the final amount of oil in the fruit was similar to that in 2021 for all treatments. The beginning of oil accumulation took place at the same time for all treatments, around the middle of Period II, and at a similar rate on the first days. During this period, Rainfed treatment had considerably less fruit oil than the rest, although it continued to increase. From the beginning of Period III, the differences were important between Rainfed and Traditional Rainfed, and Control and RDI, but not between each individual treatment among the two groups. The oil accumulation seasonal pattern showed different rates in the 2 years studied. During Period II, the rate (R2) was lower, and only at the beginning of Period III, when the endocarp reached its maximum hardness, did the accumulation rate (R3) increase considerably.

Seasonal pattern of oil accumulation during 2021 (a) and 2022 (b). Vertical bars represent standard error. Asterisks indicate significant differences in the dates shown (Turkey test; p < 0.05). Vertical dashed lines limit the period II of endocarp development. Blue circle: Control; green triangle: RDI; red square: Rainfed; yellow diamond: Traditional Rainfed (colour figure online)
The individual values of these accumulation rates (R2 and R3) were compared to the water stress indicators during each of the periods defined in Table 2. There was no significant relationship between the rates and the indicators obtained in Period I of endocarp development or they were too weak (data not shown). Table 4 and Fig. 5 show the optimum relationships obtained.

Relationship between a Min-SWP in Period II vs. oil accumulation in Period II (R2), b Min-SWP in Period II vs. oil accumulation in Period III (R3), c Av-SWP in Period III vs. oil accumulation in Period III (R3). Each point is a plot data. Blue circle: 2021 season. Green triangle: 2022 season (colour figure online)
R2 values were a lot lower than R3 for both years (Fig. 5). Oil accumulation during Period II (R2) showed a significant relationship with the Min-SWP for the same period, though more so in 2021 than in 2022 (Table 4). There was a slight variation of R2 data from 2022, despite the wide range of changes in the water status (Fig. 5a), resulting in worse fits (Table 4). The oil accumulation rate value during Period III (R3) showed similar values in 2021 and 2022 (Fig. 5b) with important relationships with the water status indicators (Table 4). The best fits were obtained with the Min-SWP from Period II (Table 4; Fig. 5). Although the best fit was linear (Table 4), the data suggest a constant value up to approximately −3.0 MPa for Min-SWP (Fig. 5b) or −2.0 MPa for Av-SWP (Fig. 5c). The R3 rate was also significantly related to the SI and the Av-SWP during Period III (Table 4). There were no fits that could link in one equation the indicators for Periods II and III with R3 (data not shown).
Discussion
Endocarp development
The endocarp development took place simultaneously with fruit volume increase and oil accumulation, but this did not match the growth rate when it was at its peak for the three processes considered (Figs. 2, 3, 4). The seasonal pattern of endocarp development was hardly affected by the fruit load conditions and water status of the trees (Fig. 2; Tables 1, 2, 3). The duration of Period I in this work was similar to that described by several authors for varieties and locations when there is no water stress (Sánchez-Piñero et al. (2022), 49 days, and Rapoport et al. (2013), 50 days). Only under very severe water stress conditions, i.e., Rainfed treatment in 2022, a change in endocarp hardening was noticed, with lower maximum values than the rest and a significantly shorter Period I duration (Fig. 2; Tables 1, 2). Data suggest that this could be related to water stress in pre-flowering stage (Table 3). Although there were no significant differences in this period for the years studied, the Min-SWP was more negative in the Rainfed treatment in 2022 vs. the rest of the treatments and years. Only one of the three Rainfed plots showed values much lower than the rest (−2.50 MPa Min-SWP), and the differential with the next more negative value (a RDI plot) was 1.14 MPa, indicating the values that could have caused the endocarp development alterations. Hammani et al. (2013), working with cv Picual, presented similar results, with a shortening of Period I and a smaller sclerified area in Rainfed trees under severe water stress (higher than −3.0 MPa during pre-flowering) but not under moderate stress conditions (the water status of this data is shown in Moriana et al. 2003). On the other hand, Rapoport et al. (2004) describe that, for cv Leccino, a severe water stress (more negative than −3.0 MPa of potential at dawn) at this stage caused the extension of Period I. However, the trees studied by Rapoport et al. (2004) received a rehydration period immediately after this, which was not the case in the current experiment, nor in the one by Hammani et al. (2013). This suggests that, under severe stress conditions, the endocarp retains its ability to grow, even at the beginning of the fast hardening period (Rapoport et al. 2004), and a lower pit hardness would be required to indicate incomplete sclerification (Hammani et al. 2013; this work).
Fruit growth
Period I of endocarp development has a definite impact on the final fruit size (Rapoport et al. 2004; Sánchez-Piñero et al. 2022) because water stress at this phenological stage may affect the number of cells (Costagli et al. 2003) and their size (Rapoport et al. 2004) in a permanent way. The response of fruit growth to water stress during Period I changed according to the fruit load level. During Period I, in 2021, which was a high yield year, there were significant differences in volume and a trend toward worse indicator values in the Rainfed treatment. However, there were no fruit volume differences in 2022, despite the higher water stress levels in RDI and Rainfed vs. Control during Period I (Fig. 3; Table 3). This suggests that fruit growth was more sensitive to water stress than SWP, as previously indicated in other works (Costagli et al. 2003; Gucci et al. 2019; Martin-Palomo et al. 2020). The olive tree physiology changed according to the fruit load level, with less negative water potential values and lower leaf conductance in years with a very low fruit load (Martin-Vertedor et al. 2011; Corell et al. 2016). Nevertheless, such differences in water relations were identified in Period II, when massive hardening began (Martin-Vertedor et al. 2011), but not in Period I. This absence of difference would be related to a lower sensitivity of water relations at leaf level vs. fruit development because it would then be possible to predict size changes during a loading or unloading year with the data from this Period I (Sánchez-Piñero et al. 2022). The sclerification process means a high sink of water and nutrients (Rallo and Suarez 1989; Girón et al. 2015; Trentacoste et al. 2022). So, although at a lower rate, it starts with fertilization (Rapoport et al. 2004). The presence of a lower number of fruits in the low fruit load season could ensure adequate fruit development under severe water stress at leaf level since the fruit would be a preferred sink. Trentacoste et al. (2022) also concluded that, even if the initial fruit growth is a weak sink, severe shadowing during Period I had a bigger impact on the high fruit load than on the low fruit load season and increased abortion due to competence. The water stress levels under high fruit load conditions that could impact volume in Period I would be between RDI and Rainfed values for 2021 (Table 3). Av-SWP data were similar to those suggested by Hueso et al. (2021), at −1.5 MPa. However, for Av and Min-SWP, there was a very narrow range—only 0.1 MPa difference between RDI and Rainfed values. This narrow difference in both indicators could suggest that this was actually a cumulative effect of the water stress, represented by a higher SI with a threshold between 6.8 and 16.3 MPa day. Such thresholds have to be checked in future works and with other cultivars.
The fruit growth process stopped suddenly with the beginning of the massive hardening (Period II) in the Rainfed treatment, but not for RDI (Figs. 2, 3; Tables 1, 2). RDI trees did not even reduce their growth rate at the beginning of this Period II, although the water potential values during Period I were very similar for both treatments and clearly worse than those for Control (Fig. 3; Table 3). These results suggest a cumulative effect of water stress, together with an increase in the sclerification process sink effect. The average values when the growth stopped in Rainfed and the rate changed in RDI were around 30 MPa day, with higher SI values for fruit volume decreased in RDI (data not shown). The different responses between Rainfed (stopped) and RDI (growth rate reduction) suggest a differential effect of water stress based on the time point when it occurred—for the first case, just at the beginning of Period II, and for RDI, during the second half of this period. Several studies suggested that SI values higher than 30 MPa day resulted in a fruit growth stop (Girón et al. 2015; Martín-Palomo et al. 2019). Other works also suggested a cumulative impact of water stress on fruit size when such conditions are applied at the end or during Period I and Period II (Gomez del Campo et al. 2014). Although the cell division process in the fruit has been described as not exclusive to Period I (Hammami et al. 2011; Gomez del Campo et al. 2014), the differential response between Rainfed and RDI could be considered related to the main impact of this process instead of cell expansion.
The volume reduction process was completely reversible in the RDI treatment, as different authors described using stress time points similar to those in this work from Period II (Moriana et al. 2003; Girón et al. 2015; Corell et al. 2020). However, under conditions of highly severe water stress in Period I or at the beginning of Period II, the size was clearly reduced despite rehydration (Moriana et al. 2003), or there were no significant differences but a clear trend toward being smaller (Gomez del Campo et al. 2014; Gucci et al. 2019). Such responses would support the idea that different processes in fruit development would be affected at different times, and this would affect the possible recovery of fruit size.
Oil accumulation
The oil accumulation process was more resilient to water stress than fruit growth. Accumulation started simultaneously in all treatments, with a seasonal continuous growth pattern in all data collected (Fig. 4), although at different rates (Fig. 5). This increase took place despite the complete stop of fruit growth in the Rainfed or RDI treatments (Fig. 3), and it happened when the water stress levels were more severe, i.e., Periods II and III (Table 3). This response, together with the previous ones, suggests a strong pattern of assimilate distribution, acting as the main sink in the pit hardening process, followed by the oil accumulation, which starts a faster accumulation period when the previous one ends, and, lastly, the fruit growth (Figs. 2, 3, 4). Different works suggest the relevance of the hardening process vs. fruit growth (Rapoport et al. 2004) or vegetative growth (Rallo and Suarez 1989; Hernandez-Santana et al. 2018).
The absence of a relationship between the rates of oil accumulation (R2 and R3) and indicators of water status in Period I suggests that the cell division could have an influence on the total oil accumulated, which is smaller than previous works suggested (Gomez del Campo et al. 2014). It also seems to indicate that a smaller final fruit size in Period I, and potentially at harvest, could be offset by a more efficient oil production during Periods II and III. In fact, the fruit volume reduction seen between DOY 220 and DOY 240 in both years (Fig. 3) did not have an impact on oil accumulation (Fig. 4). Different studies suggest that moderate water stress would probably improve the oil percentage in the fruit (Moriana et al. 2003; Grattan et al. 2006; Gucci et al. 2019; Lavee et al. 2007; Fernández et al. 2013; Ben-Gal et al. 2021; Corell et al. 2022). Water stress during Period II of endocarp development apparently had a closer relationship with the oil accumulation rate, both in this period (R2) and in the following one (R3) (Table 4). Garcia-Inza et al. (2014) carried out an experiment on the susceptibility of olive trees to high temperatures and suggested that the fruit development period for the cv Araujo during January–February (Southern hemisphere) had the biggest impact on oil accumulation. Although endocarp development was not accurately established (cutting with a knife), the period of that work (Garcia-Inza et al. 2014) could approximately match Period II in the current work.
The linear fit between R3, the steepest oil accumulation rate, and the water status indicators was good (Fig. 5; Table 4). But data suggested that this sensitivity would actually be linked to a threshold defined by water stress conditions more negative than −3.0 MPa of Min-SWP. The fits in Table 4 provided good results for SI and Av-SWP too, which could suggest that a lack of rehydration in the Rainfed treatments was the most influential factor on the Min-SWP indicator in this work. Oil accumulation was characterized as a sigmoid, where the period with the greatest oil accumulation rate could be simplified as a linear relationship (López-Bernal et al. 2021). López-Bernal concluded that the rate of this linear equation (R3 in the current work) would be less sensitive to water stress. Current data suggest that the reduction would be important at severe levels, proving the need to determine exactly the water stress to establish a possible oil yield reduction. Studies reporting differences in the oil harvest are related to water stress levels that are very severe and last a considerable amount of time (Moriana et al. 2003; Fernández et al. 2013; Ahumada-Orellana et al. 2017; Corell et al. 2022, among others). The yield reduction could be linked to a loss of fruit due to water stress conditions during Period I (e.g., Moriana et al. 2003). However, works using water stress only during Periods II and III also indicated that yield reduction would be mainly associated to an impact on oil accumulation, seeing that the fruit drop seems less sensitive to water stress in these periods (Corell et al. 2022). Irrigation works with oil production trees that are usually rehydrated due to rainfall or irrigation reported significant yield loss only under water stress conditions with water potentials below −3.0 MPa before rehydration (Ahumada-Orellana et al. 2017) or values close to that, but with a long duration (Fernández et al. 2013). Water stress conditions with minimum potentials and less durations seemed to have a lesser impact with hardly any significance (Moriana et al. 2003; Ben-Gal et al. 2021; Corell et al. 2022). This suggests that rehydration plays a role during Period III that would allow recovering part of the oil production, although not completely. Hueso et al. (2019) suggested that water stress would not affect oil production until values close to −2.21 MPa of SWP were reached in Period III. This threshold was greatly exceeded in this study (Table 3), and this would support the hypothesis that the current study suggests a cumulative rather than an ad hoc effect, in line with the other authors (Moriana et al. 2003; Fernández et al. 2013; Ahumada-Orellana et al. 2017; Ben-Gal et al. 2021). However, because, in the current work, the cultivar was for table use, these conclusions could be limited and not completely useful for oil cultivars.
Under no circumstances, did the difference in fruit load affect the oil accumulation period or the amounts produced in the fruit (Fig. 4); however, it did affect the relationship with the water stress indicators (Table 4). During the low fruit load year, Period II had a lower sensitivity to water stress than Period III, which, conversely, resulted in some similar relationships between both seasons (Fig. 5; Table 4). The massive pit hardening in Period II turned the fruit into a preferred sink for water over the leaves, resulting in the absence of response seen in the low fruit load year. On the other hand, during Period III, once Period II is over, the strength of the fruit as a sink could be reduced, thus making it more sensitive, even in low fruit load seasons.
Conclusion
The absence of changes in endocarp hardening patterns due to water stress conditions ensures an accurate schedule of RDI using this process. However, because experiments similar to the current one were scarce, this has to be checked in further studies with different cultivars. The optimization of deficit irrigation would also need continuous tree water status monitoring because the main yield components (fruit size and oil amount) were related to an accumulation of water trees. The fruit growth stage was the most sensitive process to water stress, being affected when the latter was around 30 MPa day. However, the fruit volume reduction could be reversed to levels equal to Control when it happened at Period II but not earlier. The oil accumulation started during Period II, and the rate was low (R2) in all treatments. This process was moderately resistant to water stress, but during the period of higher accumulation rate (R3), it was linked to the water status indicators of Periods II and III. Only in the year with a very low fruit load, was the fruit growth in Period I and the oil accumulation in Period II not related to the water stress indicators. This suggests that the sclerification process acted as a large water sink that isolated the fruit from the leaves under conditions of a very low load. All these conclusions show the need to accurately establish the process of hardening in the fruit. Although changes between cultivars, mainly because of their final use, could be expected, the value of cumulative stress would be demonstrated. Further work is needed to consider oil cultivars in similar studies and, in general, different levels and durations of water stress that permit the fitting threshold values accurately.
Data availability
We will upload the data openly, in the us repository: Idus.us.es.
References
AEMET (2023) Spanish agency of meteorology. Available online https://www.aemet.es/es/serviciosclimaticos/datosclimatologicos/. Accessed on 30 May 2023
Ahumada-Orellana LE, Ortega-Farias S, Searles PS, Retamanes JB (2017) Yield and water productivity responses to irrigation cut-off strategies after fruit set using stem water potential thresholds in a super-high density olive orchard. PlantSci. https://doi.org/10.3389/fpls.2017.01280
Ben-Gal A, Ron Y, Yermiyahu U, Zipori I, Naoum S, Dag A (2021) Evaluation of regulated deficit irrigation strategies for oil olives: a case study for two modern Israeli cultivars. Agric Water Manage 245:106577. https://doi.org/10.1016/j.agwat.2020.106577
Chalmers DJ, Wilson IB (1978) Productivity of peach trees. Tree growth and water stress in relation to fruit growth and assimilate demand. Ann Bot 42:285–294
Corell M, Pérez-López D, Martín-Palomo MJ, Centeno A, Girón I, Galindo A, Moreno MM, Moreno C, Memmi H, Torrecillas A, Moreno F, Moriana A (2016) Comparison of the water potential baseline in different locations. Usefulness for irrigation scheduling of olive orchards. Agric Water Manag 177:308–316. https://doi.org/10.1016/j.agwat.2016.08.017
Corell M, Martín-Palomo MJ, Girón I, Andreu L, Galindo A, Centeno A, Pérez-López D, Moriana A (2020) Stem water potential-based regulated deficit irrigation scheduling for olive table trees. Agric Water Manag 242:106418. https://doi.org/10.1016/j.agwat.2020.106418
Corell M, Pérez-López D, Andreu L, Recena R, Centeno A, Galindo A, Moriana A, Martín-Palomo MJ (2022) Yield response of a mature hedgerow oil olive orchard to different levels of water stress during pit hardening. Agric Water Manag 261:107374. https://doi.org/10.1016/j.agwat.2021.107374
Costagli G, Gucci R, Rapoport H (2003) Growth and development of fruits of olive ‘Frantoio’ under irrigated and rainfed conditions. J Hortic Sci Biotechnol 78(1):119–124. https://doi.org/10.1080/14620316.2003.11511577
De Sosa LL, Sánchez-Piñero M, Girón I, Corell M, Madejón E (2023) Addition of compost changed responses of soil-tree system in olive groves in relation to the irrigation strategy. Agric Water Manage 284:108328. https://doi.org/10.1016/j.agwat.2023.108328
Díaz-Espejo A, Fernández JE, Torres-Ruíz JM, Rodríguez-Domínguez CM, Pérez-Martin A, Hernández-Santana V (2018) The olive tree under water stress: fitting the pieces of response mechanisms in the crop performance puzzle. In: García-Tejero I, Martin-Zuago VH (eds) Water scarcity and sustainable agriculture in semiarid environment. Tools, strategies and challenges for woody crops. Academic Press, London, pp 439–480
Fernández JE, Pérez-Martin A, Torres-Ruiz JM, Cuevas MV, Rodriguez-Dominguez CM, Elsayed-Farag S, Morales-Sillero A, García JM, Hernandez-Santana V, Diaz-Espejo A (2013) A regulated deficit irrigation strategy for hedgerow olive orchards with high plant density. Plant Soil 372:279–295. https://doi.org/10.1002/jsfa.7828
Garcia-Inza GP, Castro DN, Hall AJ, Rousseaux MC (2014) Responses to temperature of fruit dry weight, oil concentration, and oil fatty acid composition in olive (Olea europaea L. var. ‘Arauco’). Eur J Agron 54:107–115. https://doi.org/10.1016/j.eja.2013.12.005
Girón IF, Corell M, Galindo A, Torrecillas E, Morales D, Dell’Amico J, Torrecillas A, Moreno F, Moriana A (2015) Changes in the physiological response between leaves and fruits during a moderate water stress in table olives. Agric Water Manag 148:280–286. https://doi.org/10.1016/j.agwat.2014.10.024
Girona J, Fereres E, Marsal J, Goldhamer DA, Naor A, Soriano MA (2012) Peach. In: Steduto P, Hsiao TC, Fereres E, Raes D (eds) Crop yield response to water. FAO, Rome, pp 392–409
Goldhamer DA (1999) Regulated deficit irrigation for California canning olives. Acta Hortic 474:369–372. https://doi.org/10.17660/ActaHortic.1999.474.76
Gómez del Campo M, Perez-Exposito MA, Hammami SBM, Centeno A, Rapoport HF (2014) Effect of varied summer deficit irrigation on components of olive fruit growth and development. Agric Water Manag 137:84–91. https://doi.org/10.1016/j.agwat.2014.02.009
Grattan SR, Berenguer MJ, Connell JH, Polito VS, Vossen PM (2006) Olive oil production as influenced by different quantities of applied water. Agric Water Manag 85:133–140. https://doi.org/10.1016/j.agwat.2006.04.001
Gucci R, Caruso G, Gennai C, Esposto S, Urbani S, Servili M (2019) Fruit growth, yield and oil quality changes induced by deficit irrigation at different stages of olive fruit development. Agric Water Manag 212:88–98. https://doi.org/10.1016/j.agwat.2018.08.022
Hammami SBM, Manrique T, Rapoport HF (2011) Cultivar based fruit size in olive depends on different tissue and cellular processes throughout growth. Sci Hortic 130:445–451. https://doi.org/10.1016/j.scienta.2011.07.018
Hammani SBM, Costagli G, Rapoport HF (2013) Cell and tissue dynamics of olive endocarp sclerification vary according to water availability. Physiol Plant 149:571–582. https://doi.org/10.1111/ppl.12097
Hernandez-Santana V, Fernandes RDM, Perez-Arcoiza A, Fernández JE, García JM, Diaz-Espejo A (2018) Relationships between fruit growth and oil accumulation with simulated seasonal dynamics of leaf gas exchange in the olive tree. Agric for Meteorol 256:458–469. https://doi.org/10.1016/j.agrformet.2018.03.019
Hsiao TC (1990) Measurements of plant water status. In: Steward BA, Nielsen DR (eds) Irrigation of agricultural crops. Agronomy monograph 30, chap 9. American Society of Agronomy, Madison, pp 243–279
Hueso A, Trentacoste ER, Junquera P, Gómez-Miguel V, Gómez del Campo M (2019) Differences in stem water potential during oil synthesis determine fruit characteristics and production but not vegetative growth or return bloom in an olive hedgerow orchard (cv Arbequina). Agric Water Manag 223:105589. https://doi.org/10.1016/j.agwat.2019.04.006
Hueso A, Camacho G, Gómez-del-Campo M (2021) Spring deficit irrigation promotes significant reduction on vegetative growth, flowering, fruit growth and production in hedgerow olive orchards (cv Arbequina). Agric Water Manag 248:106695. https://doi.org/10.1016/j.agwat.2020.106695
Lavee S, Hanoch E, Wodner M, Abramowitch H (2007) The effect of predetermined deficit irrigation on the performance of cv Muhasan olives (Olea europaea L.) in the eastern coastal plain of Israel. Sci Hortic 112:156–163. https://doi.org/10.1016/j.scienta.2006.12.017
López-Bernal Á, Fernandes-Silva AA, Vega VA, Hidalgo JC, León L, Testi L, Villalobos FJ (2021) A fruit growth approach to estimate oil content in olives. Eur J Agron 123:126206. https://doi.org/10.1016/j.eja.2020.126206
Lorite IJ, Cabezas JM, Ruiz-Ramos M, de la Rosa R, Soriano MA, León L, Santos C, Gabaldón-Leal C (2022) Enhancing the sustainability of Mediterranean olive groves through adaptation measures to climate change using modelling and response surfaces. Agric for Meteorol 313:108742. https://doi.org/10.1016/j.agrformet.2021.108742
Martín-Palomo MJ, Corell M, Girón I, Andreu L, Trigo E, López-Moreno YE, Torrecillas A, Centeno A, Pérez-López D, Moriana A (2019) Pattern of trunk diameter fluctuations of almond trees in deficit irrigation scheduling during the first seasons. Agric Water Manag 218:115–123. https://doi.org/10.1016/j.agwat.2019.03.033
Martin-Palomo MJ, Corell M, Girón I, Andreu L, Galindo A, Centeno A, Pérez-López D, Moriana A (2020) Absence of yield reduction after controlled water stress during preharvest period in table olive trees. Agronomy 10:258. https://doi.org/10.3390/agronomy10020258
Martin-Vertedor AI, Perez-Rodriguez JM, Prieto-Losada H, Fereres E (2011) Interactive responses to water deficits and crop load (Olea europea L cv Morisca) I.—growth and water relations. Agric Water Manag 98:941–949. https://doi.org/10.1016/j.agwat.2011.01.002
Moriana A, Orgaz F, Pastor M, Fereres E (2003) Yield responses of a mature olive orchard to water deficits. J Am Soc Hortic Sci 128:425–431. https://doi.org/10.21273/JASHS.128.3.0425
Myers BJ (1988) Water stress integral a link between short term stress and long term growth. Tree Physiol 4:315–323. https://doi.org/10.1093/treephys/4.4.315
Navas-Lopez JF, Leon L, Trestaconte ER, de la Rosa R (2019) Multi-enviroment evaluation of oil accumulation pattern parameters in olive. Plant Physiol Biochem 139:485–494. https://doi.org/10.1016/j.plaphy.2019.04.016
R Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/
Rallo L, Suarez MP (1989) Seasonal distribution of dry matter within the olive bearing limb. Adv Hortic Sci 123:1069–1075
Rapoport HF, Costagli G, Gucci R (2004) The effect of water deficit during early fruit development on olive fruit morphogenesis. J Am Soc Hortic Sci 129:121–127. https://doi.org/10.21273/JASHS.129.1.0121
Rapoport HF, Pérez-López D, Hammami SBM, Aguera J, Moriana A (2013) Fruit pit hardening: physical measurements during olive growth. Ann Appl Biol 163:200–208. https://doi.org/10.1111/aab.12046
Sánchez-Piñero M, Martin-Palomo MJ, Moriana A, Corell M, Pérez López D (2022) Endocarp Development study in full irrigated olive orchards and impact on fruits features at harvest. Plants 11:3541. https://doi.org/10.3390/plants11243541
SIAR (2023) Climatic station data in Spain. Available online http://eportal.mapa.gob.es/websiar/SeleccionParametrosMap.aspx?dst=1. Accessed on 30 May 2023
Trentacoste ER, Calvo FE, Sánchez CL, Calderón FJ, Banco AP, Lémole G (2022) Response of inflorescence structure and oil yield components to source-sink manipulation by artificial shading in olive. Theor Exp Plant Physiol 34:171–183. https://doi.org/10.1007/s40626-022-00239-z
Acknowledgements
This research was supported by the Junta de Andalucía and the Fondo Europeo de Desarrollo (FEDER) project P20_00492. Marta Sánchez-Piñero held a pre-doctoral contract from the University of Seville (VI-PPITUS). The authors wish to thank Finca La Hampa for their helpful assistance during the experiment and, in particular, Dr. Ignacio Girón.
Funding
Funding for open access publishing: Universidad de Sevilla/CBUA.
Author information
Authors and Affiliations
Contributions
MSP, DPL, and AM conducted research and conceptualization. MC and MJMP contributed to methodology and formal analysis. MSP and AM wrote the main manuscript text and NMZ, DPL and LLS prepared figures and tables. IG, EM, PCV contributed to review and editing. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Conflict of interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sánchez-Piñero, M., Corell, M., de Sosa, L.L. et al. Assessment of water stress impact on olive trees using an accurate determination of the endocarp development. Irrig Sci 42, 461–476 (2024). https://doi.org/10.1007/s00271-024-00914-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00271-024-00914-w