
Abstract
Urbanization and land cover change are significantly affecting the availability of habitats for wildlife worldwide. However, linking species persistence to large-scale habitat changes is challenging, especially when wildlife monitoring data is lacking. In China, the Eurasian otter (Lutra lutra) is a species of conservation concern and is listed as endangered as a consequence of declining populations across the country. Hong Kong is home to a small population of Eurasian otters, which are primarily restricted to a set of wetlands in the inner Deep Bay area. However, the drivers of their historical distribution changes are largely unknown. We combined otter spraint data from 2018 to 2019 with historical records of otter data spanning 1959–2018 to examine otter habitat preference and changes in their distribution, relating this with land cover changes measured by Landsat-5 and Landsat-8 satellite remote sensing in the past (1986 and 1995) and present (2018) using supervised random forest classification. We found that otters showed habitat preference for fish ponds and watercourses, and persisted in areas where fish pond area was more readily available. We also found no significant effect of the extent of land cover changes on otter persistence, suggesting that the species is resilient to some level of fish pond loss and find value in available terrestrial habitats. Our results demonstrate the utility of relating satellite remote sensing data to species distribution data over decadal time scales and highlight the importance of managing terrestrial and wetland habitats for otters and other key species in increasingly urbanized landscapes.
Similar content being viewed by others

Introduction
Wetlands, which provide critical habitats for a diverse range of species, are threatened by a range of drivers, leading to an estimated loss of 21% of global wetland area since 1700 (Fluet-Chouinard et al. 2023). This loss has significant global consequences for biodiversity, particularly in Asia, which has experienced some of the highest rates of wetland loss in recent decades (Hu et al. 2017). The primary drivers of wetland loss include conversion for cropland, forestry, cultivation, and urbanization (Fluet-Chouinard et al. 2023). These changes result in the loss of habitat for many species, as well as reductions in their abundance and range.
Eurasian otters (Lutra lutra) are highly dependent on wetlands for survival. These sub-aquatic, carnivorous mammals thrive in wetland environments due to the relatively shallow, intermittent inundation and ample prey availability (Kruuk 2006). Additionally, otters play a vital role in regulating wetland ecosystem function as key apex predators (Duplaix and Savage 2022). Despite their importance, research on Eurasian otters in Asia remains limited, with an increasing need to understand their population statuses and ecological needs, such as habitat conditions, diet, and behavior (Basnet et al. 2020). Such information is crucial for developing effective conservation strategies for Eurasian otters in Asia.
Existing research in Europe has shown that Eurasian otters exhibit preferences for different types of habitats. For instance, in narrow aquatic zones, they favor substrates composed of small rocks and vegetation cover (Kruuk 2006). A study in Scotland found that during winter, otters’ population size and distribution are largely determined by topography and access to open water, with food supply and population density playing secondary roles (Erlinge 1967). In Poland, river width positively correlated with their distribution, while the presence of buildings and river channels showed a negative correlation (Romanowski et al. 2013). Other studies have suggested that otter distribution is not affected by the percentage of vegetative cover or agricultural land use (Romanowski et al. 2013; Durbin 1998).
In Hong Kong, Eurasian otters are primarily found in wetlands of the inner Deep Bay area in northwest New Territories (AFCD 2018; Shek 2006). In the last 50 years, this area has undergone drastic changes due to increased demands for freshwater fish in the 1970s, leading to the development of many fish ponds that provided ideal habitats for otters (Young and Melville 1993; Hui and Chan 2023; McMillan et al. 2023) and are included within the Ramsar definition of wetlands (Ramsar Convention on Wetlands 2016). However, since the 1970s, nearly 40% of fish ponds and 21% of mangroves have been converted into urban and residential areas (Chum 2013). Further changes are anticipated following the release of the Northern Metropolis Development Strategy by the Hong Kong Government (HKSAR 2021), which aims to develop the Deep Bay area to accommodate an additional 1.5 million people, whilst designating conservation zones for the remaining wetlands. Given that this proposal covers the known distribution of Eurasian otters in Hong Kong, continual habitat changes and increasing human disturbance in the inner Deep Bay area could negatively affect their distribution. Eurasian otters are already listed as Near Threatened on the IUCN Red List of Threatened Species (Roos et al. 2015) and are protected by the Protection of Endangered Species of Animals and Plants Ordinance and Wild Animals Protection Ordinance in Hong Kong (AFCD 2018). Furthermore, the sensitivity of otters to habitat changes means they could act as indicator species for the overall health of wetland ecosystems in the Deep Bay area (Ruiz-Olmo and Jiménez 2009; Ali et al. 2010).
Whilst it is understood that the small population of Eurasian otters in Hong Kong are reliant on fish ponds for diet (McMillan et al. 2023), little is known about the impact of otter fish pond loss and urban expansion on otter populations over time. Therefore, the aim of this study is to explore the relationship between land cover changes and Eurasian otter persistence in the Deep Bay area of Hong Kong from 1986 to 2018. Towards this aim, we used field interviews for historical data (McMillan et al. 2019) and otter spraint data (McMillan et al. 2023), and leveraged long time-series satellite remote sensing data to (1) determine locations where otter populations had persisted or disappeared, (2) quantify changes in land cover, and (3) establish the relationship between land cover changes and otter persistence. We hypothesized that land cover change would impact otter persistence and that fish pond conversion in particular would be associated with loss of otters over time. Optical remote sensing has long been successfully used for mapping the extent, distribution and change in wetland habitats and urban areas in Asia and worldwide (Kayastha et al. 2012; Haque and Basak 2017; Kaplan et al. 2019; Aslam et al. 2023, 2024). With this method, this study will provide insights into the ability for otters to persist in wetland areas with direct or surrounding conversion pressures, thereby informing conservation strategies for Hong Kong and the wider region with projected urbanization of wetlands into the future.
Methodology
Study Area and Study Period
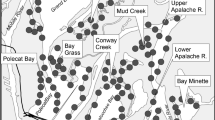
Our study area covered the Hong Kong side of the Deep Bay area (22°21’-22°32’N, 113°52’-114°08’E) (Fig. 1). The north-eastern part of the study area was Ma Tso Lung and Lok Ma Chau; the south-western part was down to Ha Pak Lai, Tuen Mun and Castle Peak. The core fish pond area in the middle of the study area included San Tin, Mai Po Nature Reserve, Tai Sang Wai, Nam Sang Wai and Fung Lok Wai where many mangroves and water courses can also be found (Chum 2013).

(A) Map illustrating location of Deep Bay area and specific locations of interest for this study; (B) Example of fish ponds in the Deep Bay area (credit: Eugene Yu Hin Yau)
We investigated land cover and otter distribution changes for the period of 1986 to 2018 because this corresponds with the criteria used by International Union of Conservation of Nature (IUCN) to assess population decline in Eurasian otter (IUCN 2012). Based on the otter occurrence data, we defined past (or historic) Eurasian otter occurrences as within 1950–2009 and current Eurasian otter occurrence as within 2010–2018 following McMillan et al. (2019). Data quality of Landsat-5 did not stabilize until 1986, therefore the study period is confined to 1986–2018 in line with regular and higher quality satellite acquisitions.
To compare land cover change between three snapshots in time, 1986 was selected as the “past” snapshot (initial state) and 2018 as the “present” snapshot (final state). 1995 was chosen because it was in the middle of the past period (1986–2009) and was the year when Mai Po and parts of inner Deep Bay were listed as Ramsar sites and there was growing recognition of the importance of wetland habitat in Hong Kong (Town Planning Board 2014). The year 2018 was chosen because Landsat imagery closest to the time when the ground truth survey was conducted was needed.
Otter Occurrence Data
McMillan et al. (2019) obtained Eurasian otters occurrence records from fishermen interviews. Interviews were conducted in fish pond areas contiguous with Mai Po Nature Reserve from May 2017 to February 2018. Fish farmers, recreational fishermen and people residing in nearby areas were interviewed to obtain Eurasian otter sighting records. Records were grouped into historical otter sightings (1950–2009) and recent otter sightings (2010–2018). Based on these records, the study area was categorized into Otter-persist areas and Otter-loss areas based on 1 km x 1 km grids. Eurasian otter home range has been documented within 7 to 40 km (Erlinge 1967; Roche et al. 1995), however these home ranges were based on linearly connected habitats (i.e. rivers, or lakes linearly connected by streams). The 1 km grid size was therefore deemed most suitable for Eurasian otter home ranges in Hong Kong, which are likely to be small relative to other populations and consist of overlapping non-linear home ranges due to their dependence on mosaicked wetland for habitat rather than rivers (McMillan et al. 2023). Otter-persist areas were places where Eurasian otter sighting records occurred in both past (1950–2009) and present (2010–2018), while Otter-loss areas were places where Eurasian otter sighting records only occurred in the past but not in present. Areas with recent otter sightings only were not included in the investigation because absence of sighting records in the past did not necessarily mean an absence of otter occurrence in that area.
When assessing the distribution of Eurasian otters, spraint survey is the most commonly used method because Eurasian otters defecate frequently at conspicuous sites, providing large number of samples and facilitating data collection (Hájková et al. 2009). Camera trap surveying is also a common method in assessing distribution. There were some camera trap records on Eurasian otters in Hong Kong from 2002 to 2006 (Shek et al. 2007). However, there was no consistent spraint or camera trap records on Eurasian otters in the last 50 years. Subsequently, spraint surveys in 2018 and 2019 have generally confirmed the findings based on the fishermen interviews (McMillan et al. 2023). However, without past data on spraints, we used spraint data to confirm preferred otter habitat for the year 2018. The 253 spraint presence points from 2018 to 2019 were reduced to 18 cluster points using the ‘point cluster’ function in QGIS 3.28.2. Points that fell within 500 m (based on the selected scale for otter home range) from each other were merged into a single point. Points aggregation was made based on the closest group being formed rather than assigning the first group within the search distance. 18 random points were generated within the Deep Bay area with 500 m minimum distance between all points using the ‘Random Points’ function in QGIS 3.28.2.
Satellite Data Source
Three cloud-free median composite Landsat images covering the study area in 1986, 1995 and 2018 were produced using Google Earth Engine (Gorelick et al. 2017). These was produced by defining a collection of images within a six-month period, applying a cloud mask which uses the pre-computed Landsat QA band from CFMask to remove cloud and cloud shadows to these images (Foga et al. 2017), and calculating the median for each pixel after cloud masking. Respective composite dates for 1986, 1995 and 2018 were 23/10/1986 to 23/04/1987, 23/10/1995 to 23/04/1996 and 23/10/2017 to 23/04/2018. Composite images from 1986 to 1995 were derived from Landsat 5 and the composite image for 2018 was from Landsat 8. Level 2, Collection 2, Tier 1 atmospherically corrected surface reflectance scenes (U.S. Geological Survey) from both platforms were used. Landsat 5 Thematic Mapper I satellite carries Thematic Mapper sensor while Landsat 8 satellite carries the Operational Land Imager (OLI) and the Thermal Infrared Sensor (TIRS). Both sensors have 30 m resolution, but OLI bands are spectrally narrower than the corresponding TM bands (Table 1). Regardless of Landsat platform and sensor, all Tier 1 Landsat data available on Google Earth Engine is considered inter-calibrated and consistent across the whole Landsat collection, regardless of sensor (Gorelick et al. 2017; U.S. Geological Survey). Therefore, classification outputs for assessing land cover change over time are comparable.
Supervised Land Cover Classification
The random forest machine learning algorithm was used to identify different land covers in the Landsat images according to their spectral signatures using the bands listed in Table 1 in Google Earth Engine (Gorelick et al. 2017). The random forest algorithm is an ensemble decision tree classifier and is widely used in remote sensing classification tasks such as land cover mapping due to its non-parametric approach and insensitivity to overfitting (Breiman 2001; Rodriguez-Galiano et al. 2012; Belgiu and Drăguţ 2016; Amini et al. 2022). Optimal models were produced using 500 trees, 2 variables at each split and a bag fraction of 0.5.
Five training classes were identified: fish pond, vegetation, water course, urban and other. Fish ponds were assumed to be water bodies that are actively managed and contained sufficient fish resources. Inter-tidal shrimp ponds (known locally as gei wai) were included with fish pond because: (1) Eurasian otters can use almost any kind of water bodies and forage in both fresh water and marine ecosystems (Kruuk 2006); (2) the number of active gei wai in Hong Kong was limited; and (3) gei wai could not be distinguished from fish pond on Landsat images by human visualization. Vegetation included mangroves, shrubs and trees. Water course included natural and modified water channels. Urban included buildings, dwellings, car parks and roads. Other referred to areas that could not attributed to any of the other four classes, which included bare soil and rock in rural areas and bare ground in areas under conversion in urban areas. Separate training and testing data was created for 1986, 1995 and 2018. 300 training points were produced through simple random sampling for each class for each year and then split using an 80:20 ratio to assemble training data for building the model and test data to evaluate the accuracy of the output, respectively. To facilitate identifying different land cover classes, we used the Color Infrared band combination (4, 3, 2 for 1986 and 1995 Landsat 5 composite images; 5, 4, 3 for the 2018 Landsat 8 composite image; Table 1) to best distinguish between vegetated areas in red, urban areas in white and watercourse and fishponds as dark areas. Fish ponds could then be separated from water courses by shape, whereby fish ponds are typically clustered together in a regular shape of rounded squares and rectangles, and water courses are channels or large and/or irregularly shaped water bodies. We checked the validity of these ground points using finer spatial resolution RGB imagery from Google Earth Pro, where imagery from equivalent years of the study area were available.
Accuracy Assessment
Accuracy assessment is important for assessing the quality of the classification. First, the accuracy of human visualization on satellite images was validated, i.e. how good the authors were at identifying different land cover types from satellite images (Supplementary Table 1). To obtain ground truth data for this exercise, ground surveys were conducted from January to March 2019. The survey covered main clusters of fish ponds in the inner Deep Bay area, including Fung Lok Wai, Nam Sang Wai, Tai Sang Wai, San Tin, Lok Ma Chau and Ma Tso Lung (Fig. 1). During the ground truth surveys, we recorded the positions of fish ponds, vegetative cover, mangrove area, water courses, buildings, dwellings and other habitats relevant to our training ROIs on satellite imagery from Google Earth Pro (Supplementary Fig. 1). This ground truth data was used to create reference ROIs on the 2018 Landsat image. We assumed there was no observable land cover change in the study area from 2017 to 2019, consistent with our ground observations during on-going field research during that period for other purposes.
Second, the accuracy of the random forest classification algorithm in replicating human visualization was assessed, i.e. how well the classified image represented human visualization of different land cover types on satellite images. A confusion matrix was produced for 1986, 1995 and 2018 by comparing the user-defined class against the mapped class for the test data points (Supplementary Tables 2, 3 and 4). The confusion matrices provided four different accuracy results: overall accuracy, producer accuracy, user accuracy and kappa coefficient. Producer accuracy measures the probability of a pixel classified correctly per referenced class while user accuracy measures the probability of a pixel classified correctly per land cover classes. Kappa coefficient measures how well the classified image agrees with the referenced data. Overall accuracy measures probability of a pixel classified correctly in the error matrix.
Errors due to misclassification in the maps can bias estimates of class area extent. Using the confusion matrices for each classification, we produced misclassification bias-adjusted area estimates for corresponding classifications using the method documented by Olofsson et al. (2013, 2014). The adjusted areas and uncertainty bounds are shown in Supplementary Tables 5, 6 and 7.
Land Cover Change Analysis
Bias-corrected change in land cover area was calculated for the wider Deep Bay area for 1986 to 1995 and 1995 to 2018 by subtracting the percentage area cover for 1986 from 1995, and 2018 from 1995. To analyze land cover changes within Otter-loss and Otter-persist grids between these time periods, classification images were clipped using the 1 × 1 km grids and bias-corrected changes in area were calculated for Otter-loss area and Otter-persist area for 1988, 1995 and 2018 based on the corresponding confusion matrix for each year. Bias-corrected change in land cover area was then calculated in the same way as outlined for the wider Deep Bay area.
Logistic Regression Analysis
Logistic regression was used to test our hypothesis that the extent of land cover change would be higher in Otter-loss area than in Otter-persist area. On the otter persistence grid layer, there were 18 grids of Otter-persist records and 17 grids of Otter-loss records. After overlaying 1995 and 2018 classification images with the otter persistence grids, each grid was clipped out such that each was equivalent to one sample. We defined Otter-persist area = 1 and Otter-loss area = 0, which represented the dependent variable of otter persistence. Otter persistence data in this study were binary variables, so logistic regression was used to test for the relationship between extent of habitat change and otter persistence. Fourteen logistic regression models were run with otter persistence as the dependent variable (Supplementary Table 9). Firstly, to test for preferred habitat availability, we tested for significant differences in the initial extent of each land cover class in 1995 between Otter-loss and Otter-persist areas. Secondly, to test whether changes in land cover between 1995 and 2018 had impacted otter persistence after the Ramsar declaration, we tested for significant differences in the extent of change in land cover area from 1995 to 2018 between Otter-loss and Otter-persist areas. Thirdly, we tested for significant differences in interactions between the initial land cover area extent in 1995 and change in land cover extent from 1995 to 2018 for Otter-loss and Otter-persist areas. Finally, tests for significant differences between otter spraint presence and land cover extent in 2018 were conducted (Supplementary Table 10). All statistical tests were performed using the base packages provided by R (version 4.3.0).
Results
Land Cover Classification Study Area
In both 1995 and 2018 imagery (Fig. 2), fish ponds (light blue) were mainly clustered at the inner most corner of Deep Bay (i.e. Tai Sang Wai, Nam Sang Wai, Fung Lok Wai) and extended upwards to Lok Ma Chau. Both years showed generally similar distributions of urbanicity (orange) on the Hong Kong side of Deep Bay, showing a reduced rate of urban development in the study area during this time period. Vegetation showed increased coverage in 2018 compared to 1995, expanding particularly into regions classified as ‘other’ as vegetation has established on exposed soil after landslides or wildfires.

(A) 1995 composite Landsat-5 image used for classification. Bands 4, 3 and 2 are presented to show clear differences in water bodies, urban areas and vegetation; (B) 2018 composite Landsat-8 image used for classification. Bands 5, 4 and 3 are presented to show clear differences in water bodies, urban areas and vegetation; (C) 1995 land cover classification; (D) 2018 land cover classification. Classifications images are overlaid with otter persistence grids. Composite Color Infrared images (A) and (B) show slightly different colorations due to spectral differences in the infrared, red and green bands between Landsat-5 and Landsat-8 (Table 1)
Accuracy Assessment
The error matrices showed four accuracy measurements (Supplementary Tables 1 to 4). Overall accuracy and Kappa coefficient (k) of human visualisation on satellite images were 95.8% and 0.879 respectively (Supplementary Table 1). 1986 classification (overall accuracy = 87.8%, k = 0.85; Supplementary Table 2), 1995 classification (overall accuracy = 86.4%, k = 0.83; Supplementary Table 3) and 2018 classification (overall accuracy = 82.2%, k = 0.82; Supplementary Table 4) also showed good accuracy results.
Land Cover Change Analysis
The direction of trends of bias-corrected land cover change for Otter-loss areas, Otter-persist areas and the wider Deep Bay area are broadly the same, but to differing extents (Fig. 3; Table 2, Supplementary Table 8). Across the entire Deep Bay area, there was an overall decrease in fish pond area from 1986 to 2018 (Fig. 3; Table 2). Fish pond area decreased by 51.4% from 1986 to 1995 but then increased by 37.7% from 1995 to 2018. Similarly, areas classified as water courses showed a decrease in area of 29.7% from 1986 to 1995 but an increase of 4.6% from 1995 to 2018. The proportion of vegetated areas continued to increase over time, with an increase of 10.1% from 1986 to 1995 and 14.3% from 1995 to 2018. This was contrasted by a continued reduction in the proportion of ‘other’ areas over time, with a decrease of 8.4% from 1986 to 1995 and a further decrease of 5.9% from 1995 to 2018. Urban areas increased in proportion by 21.3% from 1986 to 1995 but showed a decrease of 9.0% from 1995 to 2018.
From 1995 to 2018, fish pond area increased by 55.5% in Otter-loss areas but decreased by 8.4% in Otter-persist areas; urban area decreased by 21.4% in Otter-loss areas but decreased by 23.7% in Otter-persist areas; vegetation area increased by just 2.5% in Otter-loss areas but increased by 16.1% in Otter-persist areas; water course areas decreased by 6.5% in Otter-loss areas but increased by 50.2% in Otter-persist areas; and other areas decreased by 23.9% in Otter-loss areas but decreased by 10.0% in Otter-persist areas (Fig. 3). Within otter persistence zones, a greater area of fishpond was found in Otter-persist areas compared to Otter-loss areas in 1986, 1995 and 2018 (Fig. 4, Supplementary Table 8). Prior to 1995, percentage decrease of fish pond was higher in Otter-loss area (45.1%) than in Otter-persist area (17.8%) (Fig. 3).

Percentage change of different land cover types in Otter-persist, Otter-loss and whole study area from 1986–1995 and 1995–2018. Error bars represent the mean bias-corrected user classification error for that land cover class between classification years

Comparison of bias-corrected class estimates for the fish pond land cover class in Otter-persist and Otter-loss areas for 1986, 1995 and 2018. Error bars represent the bias-corrected user classification error
Logistic Regression Analysis
For logistic regressions using otter spraint data, otter spraints were significantly more likely to be found in areas with a high extent of fishpond (AIC = 26.1, Nagelkerke’s R2 = 0.72, df = 34, p = 0.002) and watercourses (AIC = 28.8, Nagelkerke’s R2 = 0.67, df = 34, p = 0.001) in 2018. Otter spraints were significantly less likely to be found in areas with a high urban area extent (AIC = 48.4, Nagelkerke’s R2 = 0.19, df = 34, p = 0.04) in 2018 (Supplementary Table 10).
For all logistic regressions using otter persistence data, no significant differences were found in land cover extent in 1995 between Otter-loss and Otter-persist zones. Furthermore, no significant differences were found in land cover change from 1995 to 2018 between Otter-loss and Otter-persist zones. There were no significant differences in interaction effects between initial land cover extent in 1995 and land cover change from 1995 to 2018 between Otter-loss and Otter-persist areas (Supplementary Table 9).
Discussion
Overall, this study showed that habitat change was widespread across the Deep Bay area from 1986 to 2018. We anticipated that loss of fish pond habitat would correlate with loss of otters, but we actually found that there was no significant difference in the loss of fish pond between Otter-loss and Otter-persist areas. However, we did find that habitat preference for otters in 2018 was for fish ponds and water courses (Supplementary Table 10), and the amount of fish pond area was consistently higher in Otter-persist areas than Otter-loss areas throughout the period of 1986 to 2018 (Fig. 4). These results partially align with our expectations, since otters were more likely to persist in areas where greater amount of fish pond was available. Together, these findings emphasize the complexity of land-use change impacts on urban carnivores and likely reflects interactions between scale, spatial configuration of habitats, and management.
From 1986 to 1995, the trends of land cover change were broadly similar in direction for Otter-loss and Otter-persist areas compared to the wider Deep Bay area (Fig. 3). In the 1980s, there was little attention to wetland conservation in the Deep Bay area. Even agricultural land could be used for non-agricultural purposes as long as no structure was built (Town Planning Board 2014). Coupled with cheaper fish imported from China and reduced demand for local fish, many fish ponds were filled in and turned to bare ground for container storage or sold to developers (Boulanger et al. 2008; HKBWS 2019). These influences likely explain the moderate percentage decrease in fish pond area and large percentage increase in ‘other’ area (which includes bare ground and bare soil) from 1986 to 1995 (Fig. 3).
Further investigation into the extent of land cover change from 1995 to 2018 shed more light on the relationship between extent of habitat change and otter persistence. No significant differences were found in the extent of change in land cover for urban, vegetation, water course or ‘other’ land cover classes between Otter-loss and Otter-persist areas (Supplementary Table 9). This was contradictory to our hypothesis and previous findings which showed that otters do not prefer unvegetated or urbanized areas because they do not provide the necessary water or food resources and have high levels of human or dog disturbance (Kruuk 2006). Firstly, this could be a product of misclassification from the random forest models, where fish pond and water courses were most commonly confused in land cover prediction because they are spectrally similar (Supplementary Tables 2, 3 and 4). It might be that otters were more easily sighted near human-dominated land cover types, such as urban or ‘other’ land cover which is an indicator of construction, development and the filling in of fish ponds, which was a potential bias from fishermen interview records. Alternatively, it may be that the designation of 1 × 1 km grids to denote otter persistence was inappropriate to describe the real home range of Lutra lutra and undermines the importance of the inner Deep Bay wetland system working as a whole to provide habitat and resources to support this population. On the other hand, these results could indicate that otters were able to persist despite neighboring land conversion, so long as remaining fish pond area was sufficient to provide food and water resources to sustain the otter population. This is supported by the greater area of fish pond available in Otter-persist zones across the study period (Fig. 4, Supplementary Table 8). Previous work has indicated that Eurasian otters perform better in terms of mortality and breeding success in areas where sufficient water bodies with high prey availability were found (Ruiz-Olmo and Jiménez 2009). Another study in a Mediterranean area also found that water bodies and food supply were key factors in Eurasian otter distribution range (Prenda et al. 2001). Therefore, it is possible that with enough fish pond area, size, depth, connectivity between ponds, adequate water quality and fish resources in the Mai Po Wetland area, the extent of habitat change had less influence over Eurasian otter persistence.
Since the allocation of many fish ponds in the inner Deep Bay area as Ramsar sites in 1995, fish pond area has overall increased, urban growth has stabilized and according to fisherman surveys, otters have been able to persist across the Deep Bay area. This study therefore offers evidence of the success of the Ramsar sites designation towards otter persistence in the Deep Bay area by maintaining conservation zones and reducing human disturbance levels. However, the Northern Metropolis Development Strategy (HKSAR 2021) has the potential to dramatically alter the Deep Bay landscape and transform the ecosystem. The Strategy includes plans for intensive development of homes and industry but also wetland conservation parks with active management for the species that live there (see Fig. 1 from McMillan et al. 2023). Whilst the Strategy clearly shows attempts to reduce encroachment of development on existing and proposed conservation zones, any neighboring development will inevitably apply pressure to existing habitat quality and connectivity. Anthropogenic pressure resulting from the development has the potential to impact the integrity of the wetland system, reducing the quality and quantity of resources required for otter foraging, breeding, resting, and movement, and ultimately the carrying capacity of the area. Reduced prey availability is of particular concern for a food-limited species (Kruuk 1995; Ruiz-Olmo et al. 2001) and could in turn reduce breeding success. It is therefore vital for management to ensure that high connectivity is sustained between fish ponds and wetland conservation zone to ensure adequate carrying capacity for Eurasian otters in the future, with all stakeholders involved as development plans are implemented (Hui and Chan 2023). We recommend continued work towards monitoring otter population distribution and numbers using spraint surveys and camera traps so that the success of the existing and newly-established protected wetland zones in supporting otter persistence can be assessed.
Overall, we demonstrate how historical ecological data can be related to long-term remote sensing data to tease out dynamic relationships between habitat change and species persistence. Through the integration of local ecological knowledge (McMillan et al. 2019) and satellite remote sensing data, we emphasize the importance of fish pond for Eurasian otter in Hong Kong. Given the small population size and conservation importance of the species (McMillan et al. 2023), priority should be given to ensuring the persistence of strong connected network of high-quality habitats for otters and the diversity of other species that rely on Hong Kong’s wetlands, particularly with the vulnerability of otters to projected climate change (Cianfrani et al. 2018). These priorities also apply to wetlands globally with projected widespread wetland loss due to encroachment of urbanization, sea level rise and pressure of climate change (Sofaer et al. 2016; Birch et al. 2022), even for Ramsar-designated sites (Xi et al. 2021).
Data Availability
All scripts for data production, processing and analysis are available via the following links on Figshare: https://figshare.com/s/5e155cb1e24a8158ef24; https://figshare.com/s/a7a7e09b08e012939d78; https://figshare.com/s/64b070a8d37674763cd5; https://figshare.com/s/9eabd220cf1425e950a5. Otter spraint locations are not publicly available in order to maintain privacy around specific otter locations to prevent hunting issues.
References
Agriculture Fisheries and Conservation Department (AFCD) (2018) Terrestrial mammals of Hong Kong. https://www.afcd.gov.hk/english/conservation/hkbiodiversity/speciesgroup/speciesgroup_mammals.html
Ali H, Saleem R, Qamer FM, Khan WA, Abbas S, Gunasekara K, Hazarika M, Ahmed MS, Akhtar M (2010) Habitat evaluation of smooth-coated otter (Lutrogale perspicillata) in indus plains of Pakistan using remote sensing and GIS. International Archives of Photogrammetry, Remote Sensing and Spatial Information Science 38:127–132
Amini S, Saber M, Rabiei-Dastjerdi H, Homayouni S (2022) Urban land use and land cover change analysis using random forest classification of landsat time series. Remote Sens 14(11):2654. https://doi.org/10.3390/rs14112654
Aslam RW, Shu H, Yaseen A, Sajjad A, Abidin SZU (2023) Identification of time-varying wetlands neglected in Pakistan through remote sensing techniques. Environ Sci Pollut Res 1–14
Aslam RW, Shu H, Javid K, Pervaiz S, Mustafa F, Raza D, Ahmed B, Quddoos A, Al-Ahmadi S, Hatamleh WA (2024) Wetland identification through remote sensing: insights into wetness, greenness, turbidity, temperature, and changing landscapes. Big Data Res 35:100416
Basnet A, Ghimire P, Timilsina YP, Bist BS (2020) Otter Research in Asia: Trends, biases and future directions. Global Ecol Conserv 24:e01391. https://doi.org/10.1016/j.gecco.2020.e01391
Belgiu M, Drăguţ L (2016) Random forest in remote sensing: a review of applications and future directions. ISPRS J Photogrammetry Remote Sens 114:24–31. https://doi.org/10.1016/j.isprsjprs.2016.01.011
Birch WS, Drescher M, Pittman J, Rooney RC (2022) Trends and predictors of wetland conversion in urbanizing environments. J Environ Manage 310:114723. https://doi.org/10.1016/j.jenvman.2022.114723
Boulanger K, Demott R, Nikitas D, Patchel B (2008) The market viability of Eco-fish in Hong Kong, unpublished IQP report. WPI, Worcester
Breiman L (2001) Random forests. Mach Learn 45(1):5–32. https://doi.org/10.1023/A:1010933404324
Chum KF (2013) Wetland Loss in Deep Bay in Recent Decades (Master’s thesis). Hong Kong: The University of Hong Kong
Cianfrani C, Broennimann O, Loy A, Guisan A (2018) More than range exposure: global otter vulnerability to climate change. Biol Conserv 221:103–113. https://doi.org/10.1016/j.biocon.2018.02.031
Duplaix N, Savage M (2022) The global otter conservation strategy. eScholarship, University of California
Durbin LS (1998) Habitat selection by five otters Lutra lutra in rivers of northern Scotland. J Zool 245:85–92. https://doi.org/10.1111/j.1469-7998.1998.tb00075.x
Erlinge S (1967) Home Range of the Otter Lutra lutra L. South Swed Oikos 18:186–209. https://doi.org/10.2307/3565098
Fluet-Chouinard E, Stocker BD, Zhang Z, Malhotra A, Melton JR, Poulter B, Kaplan JO, Goldewijk KK, Siebert S, Minayeva T, Hugelius G (2023) Extensive global wetland loss over the past three centuries. Nature 614(7947):281–286. https://doi.org/10.1038/s41586-022-05572-6
Foga S, Scaramuzza PL, Guo S, Zhu Z, Dilley RD, Beckmann T, Schmidt GL, Dwyer JL, Joseph Hughes M, Laue B (2017) Cloud detection algorithm comparison and validation for operational landsat data products. Remote Sens Environ 194:379–390. https://doi.org/10.1016/j.rse.2017.03.026
Gorelick N, Hancher M, Dixon M, Ilyushchenko S, Thau D, Moore R (2017) Google Earth Engine: planetary-scale geospatial analysis for everyone. Remote Sens Environ 202:18–27
Hájková P, Zemanová B, Roche K, Hájek B (2009) An evaluation of field and noninvasive genetic methods for estimating eurasian otter population size. Conserv Genet 10:1667–1681. https://doi.org/10.1007/s10592-008-9745-4
Haque MI, Basak R (2017) Land cover change detection using GIS and remote sensing techniques: a spatio-temporal study on Tanguar Haor, Sunamganj, Bangladesh. Egypt J Remote Sens Space Sci 20:251–263. https://doi.org/10.1016/j.ejrs.2016.12.003
Hong Kong Bird Watching Society (HKBWS) (2019) Deep Bay in a Minute. https://cms.hkbws.org.hk/cms/join-us-tw/zh-tw/project-tw/monitor-tw/other-monitor-tw/knowing-deep-bay (accessed 14/05/2023)
Hong Kong Special Administrative Region (HKSAR) Government (2021) Northern Metropolis Development Strategy Report. https://www.policyaddress.gov.hk/2021/eng/pdf/publications/Northern/Northern-Metropolis-Development-Strategy-Report.pdf (accessed 14/05/2023)
Hu S, Niu Z, Chen Y, Li L, Zhang H (2017) Global wetlands: potential distribution, wetland loss, and status. Sci Total Environ 586:319–327. https://doi.org/10.1016/j.scitotenv.2017.02.001
Hui M, Chan B (2023) Analysis of a 131-year longitudinal dataset of the eurasian otter Lutra lutra in Hong Kong: implications for conservation. Oryx 1–9. https://doi.org/10.1017/S0030605323001163
IUCN (2012) IUCN Red List categories and criteria. Version 3.1. Second edition. Gland, Switzerland and Cambridge, UK. IUCN. iv + 32pp
Kaplan G, Avdan ZY, Avdan U (2019) Mapping and monitoring wetland dynamics using thermal, optical, and SAR remote sensing data. Wetlands management: Assessing risk and sustainable solutions 87. https://doi.org/10.5772/intechopen.80264
Kayastha N, Thomas V, Galbraith J, Banskota A (2012) Monitoring wetland change using inter-annual landsat time-series data. Wetlands 32:1149–1162
Kruuk H (1995) Wild otters: predation and populations. OUP Oxford
Kruuk H (2006) Otters: Ecology, Behaviour and Conservation. Oxford University Press, New York
McMillan SE, Wong TC, Hau BCH, Bonebrake TC (2019) Fish farmers highlight opportunities and warnings for urban Carnivore conservation. Conserv Sci Pract 1:e79. https://doi.org/10.1111/csp2.79
McMillan SE, Wong ATC, Tang SSY, Yau EYH, Gomersall T, Wong PY, Vu AKH, Sin SYW, Hau BC, Bonebrake TC (2023) Spraints demonstrate small population size and reliance on fishponds for eurasian otter (Lutra lutra) in Hong Kong. Conserv Sci Pract 5(1):e12851. https://doi.org/10.1111/csp2.12851
Olofsson P, Foody GM, Stehman SV, Woodcock CE (2013) Making better use of accuracy data in land change studies: estimating accuracy and area and quantifying uncertainty using stratified estimation. Remote Sens Environ 129:122–131. https://doi.org/10.1016/j.rse.2012.10.031
Olofsson P, Foody GM, Herold M, Stehman SV, Woodcock CE, Wulder MA (2014) Good practices for estimating area and assessing accuracy of land change. Remote Sens Environ 148:42–57. https://doi.org/10.1016/j.rse.2014.02.015
Prenda J, Lopez-Nieves P, Bravo R (2001) Conservation of otter (Lutra lutra) in a Mediterranean area: the importance of habitat quality and temporal variation in water availability. Aquat Conservation: Mar Freshw Ecosyst 11:343–355. https://doi.org/10.1002/aqc.454
Ramsar Convention on Wetlands (2016) An introduction to the Convention on wetlands (previously the Ramsar Convention Manual). Ramsar Convention Secretariat, Gland, Switzerland
Roche K, Harris R, Warrington S, Copp GH (1995) Home range and diet of re-introduced European otters Lutra lutra (L.) in Hertfordshire rivers. Aquat Conservation: Mar Freshw Ecosyst 5(2):87–96
Rodriguez-Galiano VF, Ghimire B, Rogan J, Chica-Olmo M, Rigol-Sanchez JP (2012) An assessment of the effectiveness of a random forest classifier for land-cover classification. ISPRS J Photogrammetry Remote Sens 67:93–104
Romanowski J, Brzeziński M, Zmihorski M (2013) Habitat correlates of the eurasian otter Lutra lutra recolonizing Central Poland. Acta Theriol 58:49–155. https://doi.org/10.1007/s13364-012-0107-8
Roos A, Loy A, de Silva P, Hajkova P, Zemanová B (2015) Lutra lutra. IUCN Red List Threatened Species 2015:eT12419A21935287
Ruiz-Olmo J, Jiménez J (2009) Diet diversity and breeding of top predators are determined by habitat stability and structure: a case study with the eurasian otter (Lutra lutra L). Eur J Wildl Res 55:133–144. https://doi.org/10.1007/s10344-008-0226-3
Ruiz-Olmo J, López-Martín JM, Palazón S (2001) The influence of fish abundance on the otter (Lutra lutra) populations in Iberian Mediterranean habitats. J Zool 254(3):325–336
Shek CT (2006) A Field Guide to the Terrestrial mammals of Hong Kong. Cosmos Books, Hong Kong
Shek CT, Chan SM, Wan YF (2007) Camera trap survey of Hong Kong Terrestrial mammals in 2002-06. Hong Kong Biodivers 15:1–11
Sofaer HR, Skagen SK, Barsugli JJ, Rashford BS, Reese GC, Hoeting JA, Wood AW, Noon BR (2016) Projected wetland densities under climate change: habitat loss but little geographic shift in conservation strategy. Ecol Appl 26:1677–1692. https://doi.org/10.1890/15-0750.1
Town Planning Board (2014) TPB Guidelines 12C. https://www.info.gov.hk/tpb/tc/forms/Guidelines/pg12c_c.pdf (accessed 14/05/2023)
Xi Y, Peng S, Ciais P, Chen Y (2021) Future impacts of climate change on inland Ramsar wetlands. Nat Clim Change 11:45–51. https://doi.org/10.1038/s41558-020-00942-2
Young L, Melville DS (1993) Conservation of the Deep Bay Environment. In: Morton B (ed) The Marine Biology of the South China Sea. Hong Kong University, Hong Kong, pp 211–231
Funding
Jin Wu was supported by the HKU Seed Funding for Basic Research (202011159154) and the Innovation and Technology Fund (funding support to State Key Laboratories in Hong Kong of Agrobiotechnology) of the HKSAR, China.
Author information
Authors and Affiliations
Contributions
JF conceived the study under the supervision of TCB, JW, and SM. JF, MJL and TCB wrote the manuscript. JF and MJL conducted the analysis with guidance from CKFL. JF and SM conducted the fieldwork to support analysis. Manuscript editing contributions from JW, CKFL and SM.
Corresponding author
Ethics declarations
Competing Interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fung, J., Ledger, M.J., McMillan, S. et al. Satellite Remote Sensing Shows Maintenance of Fish Pond Area Improves Persistence of Eurasian Otters in Hong Kong. Wetlands 44, 34 (2024). https://doi.org/10.1007/s13157-024-01789-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13157-024-01789-5