
Abstract
Key message
The high-quality sap flow dataset of Melaleuca styphelioides could assists local councils in assessing tree water use and aids in the development of an urban tree sustainable planting management plan
Trees have a vital part to play in urban ecosystems, offering ecological, economic, and social advantages in addition to beautifying our suburbs. Knowledge of tree water use in urban environments is crucial for facilitating urban greening when there is a perception that growing trees on the street verge on expansive soils poses a risk to pavement and buildings. Information on long-term water use by individual trees in an urban environment is essential for local councils in developing urban tree management plans for sustainable planning of trees; however, this information is scarce. Micrometeorological models have commonly been used to estimate the canopy transpiration of plants in the absence of sap flow data. However, the reliability and accuracy of these models have rarely been assessed using sap flow measurements at the field site. This study aimed to provide the water use data of four individual Australian native Melaleuca styphelioides Sm. using sap flow instruments over 25 months. Tree transpiration, estimated based on three commonly used micrometeorological models using weather parameters and tree characteristics, was assessed and compared with sap flow measurements by employing the linear regression statistical analysis. The results revealed that the modified Penman–Monteith (PM) model demonstrated the highest level of accuracy among the evaluated models, consistently yielding lower errors and providing more reliable estimates of tree water use. This suggests that this model may be more appropriate for predicting plant water use in situations where sap flow data are unavailable.
Similar content being viewed by others


Introduction
The role of tree water use in urban ecosystems is vital, as it helps to regulate water use and improve the health of the environment, the quality of life for urban residents, and the sustainability of urban communities. Accurate measurement of transpiration is crucial to better understanding plant water utilization. Transpiration denotes the quantity of water released from the plant canopy and plays a key role in controlling the amount of water in the atmosphere. It also helps in the formation of clouds, which are vital for precipitation and weather patterns. Several methods have long been used to directly measure plant transpiration in the field, such as weighing lysimeter and potometer, although all have some disadvantages. In terms of practicality, weighing lysimeters are very costly, complex, and also impractical for tall trees, while potometers cause interference with the plant as transpiration was measured from the cutoff stems, which may not adequately represent the water use of the whole plant canopy and is not suitable for routine measurement. Indirect measurement using sap flow sensors, on the contrary, enables a practical and continuous measure of plant transpiration (Smith and Allen 1996; Li et al. 2014; Sun et al. 2022a, 2022b, 2023), which provides an accurate estimation of transpiration of a single tree (Grime et al. 1995).
The sap flow measurements are usually performed to estimate water uptake by trees since nearly 99% of water absorbed by the roots is released into the air through plant transpiration (Raven et al. 1999). Many commercially available sap flow instruments rely on different theoretical techniques despite that all of them are dependent on the measurement of temperature discrepancies following a pulse of heat (Vandegehuchte and Steppe 2013). Heat pulse velocity-based sap flow instruments have been widely employed in diverse scientific research such as horticulture (Green et al. 1997), forestry (Dye 1996), hydrology (Dawson 1996), and geotechnical engineering (Li and Guo 2017; Sun et al. 2021). Not all heat pulse velocity-based sap flow instruments are capable of nocturnal sap flow measurement. For instance, sensors based on the compensation heat pulse method have limitations for low sap flow measurement, which would underestimate the actual sap flow if the heat pulse velocity is below 3 cm/h (Burgess et al. 2000). However, heat ratio method (HRM)-based sensors such as the SFM1 adopted in this study have been proven to provide reliable and accurate measurements of reverse, zero, low, and high sap flow for a wide variety of plants (Burgess et al. 2001).
Leaf area index (LAI) is another parameter widely used in model parameterization for estimating tree water use. LAI represents the projected leaf area in relation to the ground area, serving as a dimensionless parameter that describes the extent of the plant canopy. LAI is a primary element in land surface parameterizations and microclimate studies, playing an important role in ecophysiology which determines canopy–atmosphere exchanges of energy fluxes. Direct LAI measurement techniques include allometry, litter collection, and harvesting, which measure the area of all the leaves of a plant using tools such as a leaf area meter. Such methods are destructive, are time-consuming, and do not truly reflect the spatial variability of canopy architecture (Roberts et al. 1996). Hemispherical photography (i.e., gap fraction methods) as an indirect approach that derives LAI from measurements of the transmission of radiation through the canopy has been widely used (Norman and Campbell 1989). Several studies have successfully used hemispheric photography to investigate plant canopy structures (Jonckheere et al. 2004). Negrón Juárez et al. (2009) found only a 6% error between calculated LAI and values measured with a CI-110 hemispherical sensor.
Previous researchers have attempted many models to estimate the water consumption of plant canopy. Ladefoged (1963) attempted to correlate sap flow to crown diameter for estimating canopy transpiration; however, no obvious correlation was found. The relationship between sap flow and leaf area was studied by Werk et al. (1988). A weak correlation indicates that it cannot be used to infer total plant transpiration. Cermark and Kucera (1987) also unsuccessfully correlated sap flow with leaf surface area. Grier and Running (1997) estimated plant transpiration with success by correlating sap flow with sapwood area. It was concluded by Hatton and Wu (1995) that a reliable estimation of whole canopy transpiration could be achieved by correlating sap flow with leaf surface area.
This study aimed to test the following hypotheses: (1) individual Australian native M. styphelioides trees exhibit significant variations in long-term water use in an urban environment and (2) different micrometeorological models vary in their accuracy for predicting the canopy transpiration of urban trees. To test these hypotheses, this study measured the water use of four M. styphelioides trees in an urban environment over a 25-month period using sap flow instruments. In addition, three commonly used micrometeorological models were employed to estimate the transpiration of the selected trees, and the model estimates were compared with the sap flow measurements.
Materials and methods
Description of the field site and tree species
Four individual Australian native species Melaleuca styphelioides Sm., grown in the eastern suburbs of Melbourne, were selected for sap flow measurements for a period of 25 months.
Two single trees (S1, S2) were located in Bayswater Reserve (37°51′S, 145°16'E) and two groups of trees (G1, G2) were located in Ferntree Gully (37°53′S, 145°16'E). Only one tree was chosen for sap flow monitoring from a group of trees.
Figure 1a shows the single M. styphelioides (S1) grown in Bayswater Reserve. It is a tree with light brown papery bark, alternate and ovate dark green leaves crowned with a sharp point, and creamy-white bottlebrush flowers. The tree was selected because it is one of the most favored Australian native species, which tolerates smog, pollution, and saline soils and has been widely planted in parks, reserves, and on the roadsides of streets. Each tree was instrumented with a sap flowmonitoring device and was surrounded by a protective cage. To ensure continuous sap flow measurement, a specifically designed power supply system (Fig. 1b) was utilized, with a 4.2-V (1000 mA) internal battery connected to two 12 V external batteries in series. All study sites are covered by expansive clays located in a temperate climate (i.e., Zone 3) (Li and Sun 2015), characterized by pronounced seasonal variations with annual mean temperature, based on observations from 1948 to 2019, ranging between 9.6 °C and 19.7 °C. The average yearly rainfall and sunshine hours are 855 mm and 2263 h, respectively. The average yearly solar radiation is 15 MJ /m2. All weather data reported were obtained from the Meteorological Station (086104) (BOM 2021).

The single M. styphelioides (S1) grown in Bayswater Reserve a tree silhouette b external power supply system
Tree characteristics
Tree height and crown height
The tree height, crown height, average crown spread, and leaf surface area were determined using photogrammetry. It is a reliable and fast technology for measuring objects from photographs. Two photographs were taken at N-S and E-W sides of the tree using a Nikon D7000 digital camera (18–105 mm lens), while a staff was held a range pole standing close to the tree trunk. A built-in spirit-level tripod was used to minimize measurement error. The distance between the tree and the camera was determined once the image filled most of the frame. To determine the scale of the images, the AutoCAD® software was used, enabling the determination of actual tree height and crown height. The mean value calculated from two photographs was reported. A digital laser inclinometer was employed to calibrate the tree height and crown height determined by photogrammetry. The fraction of crown height to tree height, referred to as the live crown ratio (LCR), was expressed in percentage and served as a helpful indicator for tree vigor. To calculate the diameter at breast height (DBH), a tape measure was used to determine the tree’s circumference.
Average crown spread
The average crown spread is the average length of the longest distance between the outmost foliage across the crown and the longest perpendicular spread. A photograph was taken at the base of the tree using a fisheye lens to include the whole crown silhouette, which was then imported into AutoCAD® to determine the longest spread and cross-spread, allowing for the identification of the outermost foliage onsite. The measurements of the two spreads were obtained using a tape measure.
Leaf surface area
Measuring the leaf surface area of trees is a formidable task. This study introduced a method to accurately measure leaf surface area using photogrammetry. The estimated leaf surface area reported in Table 1 is the mean of four sub-samples. Typical leaves were removed at the petiole junction and attached to a scaled white paper before a photograph (Fig. 2a) was taken using a digital camera. AutoCAD® was used to duplicate the leaf outline and subsequently calculate the actual leaf surface area, as shown in Fig. 2b.

Typical leaf shape of M. styphelioides a image captured by a digital camera b duplicated leaf silhouette
Crown shape and canopy area
Using geometric solids to simulate tree crown shapes has been widely applied with success (Halverson et al. 1980; Thayer and Maeda 1985; Mcpherson and Rowntree 1988). The tree crown shape was determined using the geometric solids developed by Coder (2005). The tree canopy area was estimated from the fisheye photograph taken below the canopy.
Leaf area index (LAI)
The CI-110 (ICD, Camas, USA) was employed to measure LAI of the whole canopy of M. styphelioides due to the availability of affordable, capable for producing high-resolution fisheye images, and easy-to-use processing software. The CI-100 consists of a digital camera (resolution of 768 × 494 pixels) with an auto-leveling PENTAX fisheye lens (150°) to scan plant canopies, and a 400-mm-long arm equipped with 24 photosynthetically active radiation (PAR) sensors to measure light. The instrument only requires one scan beneath the canopy, allowing onsite measurement in either sunny or cloudy weather conditions. The CI-100 was connected to a laptop computer via a USB cable while taking measurements, allowing the whole tree canopy to be seen on the screen and the position of the image to be adjusted. A solar disc was attached to the flexible copper tubing to block sunlight from directly striking the hemispheric lens. The instrument was placed below the canopy on the north, south, east, and west sides of the tree trunk and photographs were taken and subsequently processed externally using the Plant Canopy Analysis System that differentiated the gray levels of the sky (background) from the leaves (foreground) (Jansing et al. 1999). This system allows users to select a number of azimuthal divisions and zenith angles and mask areas with interferential objects (e.g., vegetation, cars, and people) in the image. During photograph capture, the brightness and contrast were adjusted to provide better visual quality. Ten zenith angles and ten azimuthal divisions were adopted, and Otsu’s gap fraction threshold method was used (Fig. 3). LAI was calculated by the system using the gap fraction inversion procedure based on the work by Norman and Campbell (1989). The LAI reported is the average of four readings measured in different directions.

Below-canopy hemispherical photograph captured by the CI-100 on the north side of the M. styphelioides (S1) tree grown in Bayswater Reserve
Basic wood properties
The transportation of sap and nutrients from the tree root to the leaves occurs through the water-conducting sapwood. It is crucial to assess sapwood properties, including water content and thermal diffusivity, as these parameters play a vital role in estimating the rate of tree sap flow. Increment cores extracted from tree stem representing tree diameter have been widely used to estimate sapwood depth. In this study, the 2-threads Haglöf Increment borer with a length of 400 mm was employed to extract two cores on the north side at breast height. One sample was used to determine xylem radius (i.e., the sum of the thickness of sapwood and heartwood), while another one was used for thermal diffusivity determination. The coring tool was bored halfway into the tree trunk, allowing the 5.1-mm diameter intact core sample to be obtained from the extractor sleeve. The pH indicator dye (e.g., methyl orange) was smeared on the core samples immediately upon the extraction to prevent moisture loss and to differentiate heartwood from the sapwood. The yellow region across the core sample indicates sapwood and the deep red color region is heartwood (Fig. 4). The radial thicknesses of the sapwood and heartwood were determined using a digital caliper. Bark depth is calculated as the average of four measurements conducted at different locations at breast height using a bark depth gauge. The sum of sapwood depth and heartwood depth gives the xylem radius. The cross-sectional area at breast height is known as tree basal area (TBA). Dividing the sapwood area by TBA yields sapwood percentage.

Sapwood depth estimate using pH indicator dye in M. styphelioides (S1) tree grown in Bayswater Reserve
Thermal diffusivity determination
After being removed from the extractor sleeve, the core samples were wrapped with an aluminum foil and placed in twin-zip bags to minimize water loss. They were then promptly transferred to the laboratory to determine the density and thermal diffusivity of sapwood. A sharp blade was used to remove the sapwood from the intact core. The diameter was determined by averaging four different measurement positions along the core using a digital Vernier caliper. Sapwood density was obtained by dividing oven-dried weight by fresh volume. The sapwood core was weighed after drying in the oven at 103 °C for 48 h (Sun et al. 2022a).
The thermal diffusivity (k (mm2/s)) of sapwood measures the transient thermal response of sapwood to temperature change and is calculated using Eq. 1 (Marshall 1958).
where K is the thermal conductivity, \(\rho\) is the density, and \(c\) is the specific heat capacity which can be calculated following Edwards and Warwick (1984):
where \(c_{w}\) is the specific heat capacity of the wood matrix (1200 J/K/kg is adopted) and \(c_{s}\) is the specific heat capacity of water (4182 J/K/kg is used); \(w_{f}\) is the sapwood fresh weight;\(w_{d}\) is the oven-dried weight.
where \(m_{c}\) is the sapwood water content; \(\rho_{b}\) is the sapwood density; \(\rho_{s}\) is the water density; Ks is the water thermal conductivity (0.598 W/(mK) is used); and \(K_{w}\) is the thermal conductivity of dry wood matrix and is given as (Swanson 1983):
Models for predicting tree transpiration
It is commonly accepted that micrometeorological approaches can provide reasonable measurements of plant transpiration, although such methods can be difficult to apply for spaced plantings in an orchard (Edwards and Warwick 1984). Three commonly used micrometeorological models, including the modified Penman–Monteith equation (Thorpe 1978), ASCE Penman–Monteith (PM) equation (Jensen et al. 1990), and ASCE Standardized PM Equation (Allen et al. 2005), for estimating transpiration, were introduced and employed to calculate whole canopy transpiration of the single M. styphelioides (S1). The selected models have been widely used to estimate reference evapotranspiration of plants under various climatic conditions with reasonable accuracy depending on the quality of the weather data required (Caspari et al. 1993; Edwards and Warwick 1984; Pereira et al. 2006; Amazirh et al. 2017). The results were compared with sap flow measurements by a heat pulse velocity-based sap flow sensor.
Modified Penman–Monteith equation (Thorpe 1978)
The transpiration rate of an individual plant follows Thorpe (1978) which is determined by summing over the population of individual hypostomatous leaves (Green 1993), considering variables within the canopy, including net radiation (Rn), leaf boundary layer resistance (ra), and vapor pressure deficit (D), and assuming all leaves are uniformly illuminated and D does not vary through the canopy. Equation 5 provides transpiration (E) estimate for a tree which comprises a “radiation term” and a dominant environmental component, viz., “aerodynamic term” that contributes more than 70% of the total transpiration (Green 1993):
where λ is the latent heat of water (MJ /kg); \(\Delta\) is the slope of the saturation vapor pressure temperature relationship (kPa /°C); Rn is net radiation (MJ /m2); ρa is the mean air density (kg /m3); Cp is the specific heat (MJ /kg/°C) of air at constant pressure; D is the saturation vapor pressure deficit of air (kPa); rs is the bulk surface resistance (s/m); ra is the leaf boundary layer resistance (s/m) which determines how quick the energy obtained from radiation will be lost by evaporation and forced convection; \(\gamma\) is the psychrometric constant (kPa /°C); and A is the total leaf area.
ASCE-Penman–Monteith (PM) equation (Jensen et al. 1990)
Plant transpiration occurs from the evaporation of the leaf substomatal cavity governed by the stomatal resistance consisting of parallel resistances of the top and bottom sides of leaves. The ASCE-PM equation (Eq. 6) included the surface resistance to water flux (Allen et al. 1989). This equation embodied the evaporative surface of the plant canopy as a single big leaf (Raupach and Finnigan 1988) with two parameters—one of which is dependent on atmospheric physics (ra), while another one is determined by the biological behavior of the plant canopy surface (rs).
where G is the soil heat flux (MJ /m2); \(\rho_{w}\) is the density of water (Mg /m3); ea is the actual vapor pressure of the air (kPa); es is the saturation vapor pressure of air (kPa); stomatal resistance (rs) and aerodynamic resistance (ra) are calculated using Eqs. 7 and 8 (Allen et al. 1989):
where zw is the measurement height of wind (m); zh is the measurement height of air temperature and humidity (m); d is the zero plane displacement height (m); k is the von Karman’s constant; zoh is the roughness length for transferring vapor and heat (m); zom is the roughness length governing momentum transfer(m); and uz is the wind speed at height z (m/s):
where \(r_{l}\) is the leaf stomatal resistance (s/m).
ASCE standardized-PM equation (Allen et al. 2005)
The ASCE standardized-PM equation (Eq. 9) was derived from Eq. 6 by fixing the height of 0.12 m for short crop and 0.5 m for tall crop as introduced in the ASCE Manual 70 (Jensen et al. 1990). For simplicity, the value of 2.45 MJ/kg is adopted for the latent heat of water. The albedo of 0.23 is used for the reference vegetation and 70 s/m is employed for bulk surface resistance. The height of air temperature and water vapor needs to be measured 1.5–2.5 m above the ground, while the wind speed is required to be corrected to a measuring height of 2 m:
where u2 is the daily average wind speed (m/s); T is the daily average air temperature (°C); Cn and Cd are numerator constant and denominator constant for reference vegetation, respectively.
Sap flow meter
The Sap Flow Meter SFM1 (ICT International PTY Ltd., Armidale, Australia) includes a heater needle and two temperature needles that are 35 mm long tubes individually connected to the device by a color-coded cable. SFM1 has employed the theory of heat ratio method (HRM), measuring heat movements in the water-conducting xylem tissue between two symmetrically installed temperature needles following an application of a heat pulse by a line heater positioned in the midpoint of two temperature sensors. Sap velocity is subsequently derived with corrections for probe misalignment, wounding of xylem tissue that occurred at the installation stage and thermal diffusivity (Burgess et al. 2001).
In this study, one SFM1 was installed on each tree to monitor their water usage, assuming that all trees possess a symmetrical cross-section. The pre-calibration of the device prior to installation is not required. The thick tree bark was removed before installation. The logger was programmed to record sap flow data at 30-min intervals throughout the monitoring periods. Subsequently, the sap flow data in CSV format were imported into the Sap Flow Tool software to calculate the total tree water use by multiplying the sap flow velocity with the cross-sectional area of sapwood.
Estimation of canopy transpiration
Daily canopy transpiration of the two single M. styphelioides trees (S1 and S2) grown in Bayswater Reserve was estimated using three different micrometeorological models, namely modified PM (Thorpe 1978), ASCE-PM (Jensen et al. 1990), and ASCE-standardized PM (Allen et al. 2005) for a period of 10 days in January. The results were then compared with sap flow measured using HRM-based SFM1. All the meteorological parameters required by these models were extracted from the Meteorological Station (086104), which is 2.5 km away from the field monitoring sites. The average daily temperature was obtained by averaging the daily maximum and minimum temperature which was measured by wet and dry thermometers in a Stevenson Screen at a height of approximately 1.5 m above the ground. Relative humidity was calculated from differences in temperature between the two thermometers. Wind speed was measured at 10 m above the ground by a rotating cup anemometer, which was then corrected to a measuring height of 2 m. Global solar radiation was obtained using a ground-based CM-11 pyranometer manufactured by Kipp & Zonen (BOM 2021). A transit-time porometer AP4 was used to measure stomatal conductance. Four leaf samples from various branches on the north side of the tree trunk were selected to conduct stomatal conductance measurement, and the mean was adopted for the transpiration estimation.
Results
Tree information
The characteristics of the four trees are provided in Table 1. All trees are middle aged (Mcpherson and Graves 1984), with measured tree heights ranging from 8.4 m to 8.8 m. The two single trees have a similar crown height with a mean of 7.55 m, 28% higher than the crown height of the two trees in groups, which varied between 5.4 m and 5.5 m.
The significantly larger LCR was calculated for single trees, providing an indication of healthier and faster growing. This tree species has a small leaf surface area, with the measured value varying slightly from 0.5 cm2 to 0.66 cm2. The average crown spread and canopy area for all sampled trees ranged from 4.5 m to 8.0 m and varied between 15.2 m2 and 40 m2, respectively, with distant larger crowns possessed by the two single trees. The LAI of the two trees in groups ranged between 1.4 and 1.6, 24–39% smaller than the LAI measured for the two single trees.
Wood properties
Basic wood properties for the four M. styphelioides are provided in Table 2. This species has a very thick bark with measured depths ranging from 11.6 to 22.9 mm. The sapwood depth varied from 17.58 mm to 52.83 mm, with a noticeable lower depth for trees in groups compared to isolated trees. The sapwood area of different tree trunks from the same species differs significantly from 112.6 cm2 to 844.7 cm2 as a result of the variability of tree growth. The single tree (S1) has the largest TBA of 2873 cm2; almost one-third of its area is occupied by sapwood. The sapwood percentage for trees in groups ranged between 16 and 19%, 14% to 45% smaller than that calculated for the two single trees. Parameters for determining k are shown in Table 3. The k values ranging from 0.222 mm2/s to 0.295 mm2/s were calculated for the sampled trees, which lie within a reasonable threshold between 0.14 mm2/s (water at 20 °C) and 0.40 mm2/s (dry wood) (Holman 2002).
Sap flow monitoring
Daily sap flow patterns of the four trees over February 2018 are presented in Fig. 5. The single tree’s (S1) daily peak sap flow was leveled at approximately 25 L/h over the study period, significantly higher than the daily maximum sap flow ranging from 3 L/h to 3.5 L/h for the two trees in groups (G1 and G2).

Sap flow rate comparison of the four M. styphelioides trees over February 2018
The comparison of monthly sap flow volume for the four trees from September 2017 to September 2019 presented in Fig. 6 reveals similar variation patterns of water consumption, despite great differences in daily water use. The single tree (S1) had transpired 8.6 kL of water in January 2018, which is 50%, 88%, and 87% higher than the water consumed by the single tree (S2) and trees in groups (G1) and (G2), respectively, for the same period. The monthly average temperature and monthly average solar radiation plotted in Fig. 6 show that these two weather parameters are well correlated with the monthly sap flow volume for all the four trees.

Comparison of monthly water use of the four M. styphelioides trees over 25 months and monthly average temperature and monthly average solar radiation for the same period
Comparison of transpiration derived by different models with sap flow measurements
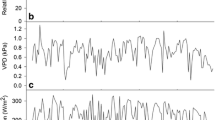
The daily average values of wind speed, solar radiation, relative humidity, and temperature, which are used to estimate canopy transpiration of the two single M. styphelioides (S1 and S2) using micrometeorological models, are shown in Fig. 7. Figure 8 compares tree transpiration estimated using three models with sap flow measurements for the two trees. Fairly similar daily sap flow patterns were observed for both the field measurements and the prediction by different models. From Fig. 8, it can be observed that sap flow estimated by the ASCE-PM model tends to be higher than the measurements for both trees. Conversely, the modified PM model yields slightly lower sap flow estimates. Interestingly, the ASCE-standardized PM model predicts transpiration that is significantly higher for S2 and lower for S1 compared to the actual measurements.

The mean daily value of various weather parameters for the study site

Sap flow measurements compared with transpiration derived from different models a S1 b S2
The linear regression statistical analysis was conducted on the data shown in Fig. 8 to assess and compare tree transpiration estimated using three models with sap flow measurements, and the results are presented in Table 4. A coefficient of determination (R2) close to 1 indicates an excellent correlation between the estimated transpiration and sap flow measurements. Figure 9 reveals that the ASCE-PM and the modified PM model achieved the highest R2 values of 0.84 and 0.83, respectively, for S1 and S2. These values signify a robust positive linear relationship. The slope is an indicator of model bias, a slope less than 1 indicates an underestimation, whereas a slope greater than 1 represents an overestimation of the true water use of trees. Error was calculated by deriving the slope from 1. Standard deviation (SD) measures the dispersion of a dataset relative to its mean. Percentage difference shows the difference in percentage between the estimated and the measured sap flow. Root mean square error (RMSE) measures how concentrated the data are around the line of best fit, and a smaller value indicates a more accurate model.

Correlation between transpiration derived from different models and sap flow measurements for the two single trees (S1 and S2) (a) modified PM (b) ASCE-PM (c) ASCE-standardized PM
Discussion
The daily sap flow for all trees displayed a “bell-shaped” pattern. It exhibited a notable increase in the early morning, followed by fluctuations around the peak during the afternoon. These variations were in response to changing light conditions and evaporative demand. Subsequently, the sap flow gradually declined and reached near-zero levels around midnight. These observations are consistent with Sun et al. (2022b), who found that the maximum daily sap flow rate of three different tree species occurred in the afternoon over a monitoring period of 12 months. The evident low rate of sap flow was observed for all the four trees on February 24, most likely due to the lowest daily solar radiation of only 3.2 MJ /m2, 84% lower than the monthly mean of 20.4 MJ /m2. The same findings were obtained by Sun and Li (2023a) that low solar radiation resulted in slow sap flow of a large Eucalypt tree.
The monthly water use of four trees showed a significant increase during the spring, with the maximum water use occurring in the early summer months. A gradual decrease in water use was observed from late summer to mid-winter before a progressive rise in the following spring months. The patterns of monthly tree water use align with the water usage trend in the same tree species, as observed by Sun et al. (2022a). The water use in the summer months (December–February) occupied 33–39% of total annual water consumption for all the sampled trees. This finding is similar to the summer water usage of a Corymbia maculata, which utilized 16 kL of water during the summer, accounting for 30% of its annual water usage (Sun and Li 2023b). An intriguing observation is that the two trees in groups (G1 and G2) managed to survive a winter month by utilizing 198 L (G1) and 193 L (G2) of water, resulting in an average daily water consumption of merely 6.6 L (G1) and 6.4 L (G2).
Tree water use can be significantly influenced by various tree characteristics, including the area of sapwood, the size of leaves and canopy, and the interception of solar radiation (Francescantonio et al. 2018; Sun and Li 2023c). Significant lesser water use by trees in groups compared to the isolated trees is mainly due to intense competition with neighboring trees for sources including water and light (Sun et al. 2022a). The canopy of the taller tree in a tree group could block out the canopy of relatively short trees and thus intercept more solar radiation, contributing to a higher transpiration rate and water use. The mean monthly water use of G1 is 6% higher than G2 at the Ferntree Gully site, although the canopy area of G2 is more than 50% larger than G1. G2 might be the vulnerable one in the group, making it less competitive for water. The mean monthly water use by S1 was 45% higher than S2 at the Bayswater site. This was most likely attributed to the substantially larger water-conducting sapwood area for S1. The differences in water use of the four trees are less likely caused by variations in the soil profile (e.g., soil water availability), as all trees were generally grown in a layer of silt loam underlain by silty clays (Carminati et al. 2016).
A fairly similar daily transpiration variation trend was observed for the field measurements and the estimate derived from three micrometeorological models, although the quantity of water extracted varied substantially daily. The ASCE-PM model consistently overestimates daily canopy transpiration relative to field measurements, with an average bias of 7.5 and 1.7% for S1 and S2, respectively. In contrast, the modified PM model yields slightly lower sap flow estimates (8.5% and 4.3% for S1 and S2, respectively). Notably, the ASCE-standardized PM model exhibits substantial variations in its predictions, with an average estimated value of 15.8% lower for S1 and 90% higher for S2 compared to the field measurements.
The ASCE-standardized PM model consistently exhibited the lowest level of accuracy among the three models, as evidenced by its highest error, mean percentage difference, and RMSE for both trees. This is mainly due to the limitations and potential shortcomings associated with the model, as it assumes an unlimited water supply for homogeneous reference crops with a specific height, which may not reflect all crops or growing environments (Pereira et al. 2006). Besides, the model assumed a constant value for the albedo of reference crops. The plant albedo varies with the color, structure, and angle of leaves and between plant species. A constant bulk surface resistance rs was also assumed, which may not accurately reflect the changing rates in leaf stomatal resistance under various weather conditions. Blonquist et al. (2010) noted that the rs may include compensation to account for any systematic bias in the ASCE-standardized PM model. The modified PM model outperforms the other two models in terms of accuracy, evidenced by its consistently lower error, mean percentage difference, and RMSE for both trees. However, there are some limitations when applying this model for plant transpiration prediction. The model requires multiple meteorological variables, which may not be readily available in all regions. Some climate parameters included in the model, such as VPD, require high-quality air temperature and relative humidity data to be available to provide accurate VPD calculation. The quality of these input data and the data consistency (e.g., missing data for a short period) can affect the accuracy of transpiration prediction (Allen et al. 2005; Sun et al. 2017). Drought stress was not observed during the study period, which might affect the evaluation performance of the modified PM model that does not consider the water stress-induced stomata closure.
Conclusions
The comparison of monthly sap flow volume for four Australian native M. styphelioides located in Melbourne, Australia, showed a similar variation trend in water use, despite marked differences in daily water consumption. The highest water demand occurred in the summer, contributing 33 to 39% of total annual water use. The single tree (S1) consumed 8.6 kL of water in a summer month, which is 88% higher than the water used by the two trees in groups (G1 and G2). The lowest transpiration occurred in winter, with monthly water use of only 198 L (G1) and 193 L (G2) for the trees in groups.
Three micrometeorological models were used to estimate the daily canopy transpiration for the two single M. styphelioides (S1 and S2) for a period of 10 days. The results were evaluated and compared with sap flow measurements using SFM1. Analysis using linear regression indicated strong positive correlations between both the modified PM and ASCE-PM models and relative changes in transpiration. The modified PM model allows the highest level of accuracy for tree transpiration predictions based on high-quality meteorological parameters. However, it should be used with caution due to its limitations and potential for inaccuracies, and appropriate adjustments should be made based on local conditions and data quality.
The tree water usage data collected from the long-term field monitoring can serve as a valuable resource for local governments, which enables them to evaluate tree water consumption and facilitate the development of efficient and cost-effective urban tree planting management plan. Arborists and botanists can also use the highly accurate transpiration model proved by this study to estimate plant water use in the absence of sap flow measurements.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Allen RG, Jensen ME, Wright JL, Burman RD (1989) Operational estimates of reference evapotranspiration. J Agron 81:650–662
Allen RG, Walter IA, Elliott RL, Howell TA, Itenfisu D, Jensen ME, Snyder RL (eds) (2005) The ASCE standardized reference evapotranspiration equation. American Society of Civil Engineers, Reston, Virginia, USA
Amazirh A, Er-Raki S, Chehbouni A, Rivalland V (2017) Modified Penman-Monteith equation for monitoring evapotranspiration of wheat crop: Relationship between the surface resistance and remotely sensed stress index. Biosyst Eng 164:68–84. https://doi.org/10.1016/j.biosystemseng.2017.09.015
Blonquist JM Jr, Allen RG, Bugbee B (2010) An evaluation of the net radiation sub-model in the ASCE standardized reference evapotranspiration equation: implications for evapotranspiration prediction. Agric Water Manag 97:1026–1038. https://doi.org/10.1016/j.agwat.2010.02.008
BOM (Bureau of Meteorology) (2021) Climate Data Online. Website: http://www.bom.gov.au/climate/data/. Accessed 19 Oct 2021
Burgess SSO, Pate JS, Adams MA, Dawson TE (2000) Seasonal water acquisition and redistribution in the Australian woody phreatophyte, Banksia prionotes. Ann Bot 85:215–224. https://doi.org/10.1006/anbo.1999.1019
Burgess SSO, Adams MA, Turner NC, Beverly CR, Ong CK, Khan AAH, Bleby TM (2001) An improved heat pulse method to measure low and reverse rates of sap flow in woody plants. Tree Physiol 21:589–598. https://doi.org/10.1093/treephys/21.9.589
Carminati A, Zarebanadkouki M, Kroener E, Ahmed MA, Holz M (2016) Biophysical rhizosphere processes affecting root water uptake. Ann Bot 118:561–571. https://doi.org/10.1093/aob/mcw113
Caspari HW, Green SR, Edwards WRN (1993) Transpiration of well-watered and water-stressed Asian pear trees as determined by lysimetry, heat-pulse, and estimated by a Penman-Monteith model. Agric for Meteorol 67:13–27
Cermark J, Kucera J (1987) Transpiration of mature stand of spruce (Picea abies (L.) Karst) as estimated by tree-trunk heat balance method. In: Forest Hydrology and Watershed Management. Proc Vanc Symp 167:311–317
Coder KD (2005) Crown shape factors & volumes. University of Georgia, Warnell School of Forestry and Natural Resources
Dawson TE (1996) Determining water use by trees and forests from isotopic, energy balance and transpiration analyses—the roles of tree size and hydraulic lift. Tree Physiol 16:263–272
Dye PJ (1996) Response of Eucalyptus grandis trees to soil water deficits. Tree Physiol 16:233–238. https://doi.org/10.1093/treephys/16.1-2.233
Edwards WRN, Warwick NWM (1984) Transpiration from a kiwifruit vine as estimated by the heat pulse technique and the Penman- Monteith equation. New Zealand J Agric Res 27:537–543
Francescantonio DD, Villagra M, Goldstein G, Campanello PI (2018) Leaf phenology and water-use patterns of canopy trees in Northern Argentinean subtropical forests. Tree Physiol 38(12):1841–1854. https://doi.org/10.1093/treephys/tpy072
Green SR (1993) Radiation balance, transpiration and photosynthesis of an isolated tree. Agric for Meteorol 64:201–221
Green SR, Clothier BE, McLeod DJ (1997) The response of sap flow in apple roots to localised irrigation. Agric Water Manag 33:63–78
Grier CC, Running SW (1977) Leaf area of mature north-western coniferous forests: relation to sit water balance. Ecology 58:893–899
Grime VL, Morison JIL, Simmonds LP (1995) Including the heat storage term in sap flow measurements with the stem heat balance method. Agric for Meteorol 74:1–25
Halverson HG, Mawson JG, Payne BR (1980) A computer program to map tree crown shadows in the urban forest. USDA Forest Service Technical Report NE-59, Broomall Pennsylvania USA
Hatton TJ, Wu HI (1995) Scaling theory to extrapolate individual tree water use to stand water use. Hydrol 9:527–540
Holman JP (2002) Heat Transfer, 9th edn. McGraw-Hill ((ISBN 978-0-07-029639-8))
Jansing E, Albert T, Chenoweth D (1999) Two-dimensional entropic segmentation. Pattern Recognit Lett 20:329–336
Jensen ME, Burman RD, Allen RG (eds) (1990) Evaporation and irrigation water requirements ASCE Manuals and Reports on Engineering Practices No. 70. American Society of Civil Engineers, New York
Jonckheere I, Fleck S, Nackaerts K, Muysa B, Coppin P, Weiss M, Baret F (2004) Review of methods for in situ leaf area index determination: Part I. Theories, sensors and hemispherical photography. Agric for Meteorol 121:19–35. https://doi.org/10.1016/j.agrformet.2003.08.027
Ladefoged K (1963) Transpiration of forest trees in closed stand. Physiol Plant 16:378–414
Li J, Sun X (2015) Evaluation of changes of Thornthwaite Moisture Index in Victoria. Aust Geomech J 50:39–49
Li J, Guo L (2017) Field investigation and numerical analysis of residential building damaged by expansive soil movement caused by tree root drying. J Perform Constr Facil 31:1–10. https://doi.org/10.1061/(ASCE)CF.1943-5509.0000908
Li J, Zhou Y, Guo L, Tokhi H (2014) The establishment of a field site for reactive soil and tree monitoring in Melbourne. Aust Geomech J 49:63–72
Marshall DC (1958) Measurement of sap flow in conifers by heat transport. Plant Physiol 33:385–396
McPherson EG, Graves GH (1984) Ornamental and shade trees for Utah. Utah State University cooperative extension service, Utah State University
Mcpherson EG, Rowntree RA (1988) Geometric solids for simulation of tree crowns. Landsc Urban Plan 15:79–83
Negrón Juárez RI, da Rocha HR, e Figueira AMS, Goulden ML, Miller SD (2009) An improved estimate of leaf area index based on the histogram analysis of hemispherical photographs. Agric for Meteorol 149:920–928. https://doi.org/10.1016/j.agrformet.2008.11.012
Norman JM, Campbell GS (1989) Canopy Structure. In: Pearcy RW, Ehlesinger J, Mooney HA, Rundel PW (eds) Plant Physiological Ecology. Field Methods and Instrumentation. Chapman and Hall, London
Pereira AR, Green S, Villa Nova NA (2006) Penman-Monteith reference evapotranspiration adapted to estimate irrigated tree transpiration. Agric Water Manag 83:153–161. https://doi.org/10.1016/j.agwat.2005.11.004
Raupach MR, Finnigan JJ (1988) Single-layer models of evaporation from plant canopies are incorrect but useful, whereas multilayer models are correct but useless: Discuss. Aust J Plant Physiol 15:706–716
Raven PH, Evert RF, Eichhorn SE (1999) Biology of Plants, 6th edn. W.H. Freeman and Company, New York USA
Roberts J, Cabral OM, da Costa JP, Mcwillian A, de Sa´ T (1996) An overview of the leaf area index and physiological measurements during ABRACOS. In: Nobre C, Roberts J, Victo´ ria R, Gash J (eds) Amazon Deforestation and Climate. J Wiley & Sons, Chichester UK
Smith DM, Allen SJ (1996) Measurement of sap flow in plant stems. J Exp Bot 47:1833–1844
Sun X, Li J (2023a) Case study of ground subsidence caused by the drying effect of a group of Australian native eucalypts. J Perform Constr Facil 37(5):1–11. https://doi.org/10.1061/JPCFEV.CFENG-4254
Sun X, Li J (2023b) A Parametric study of the effect of trees on residential footing design on expansive soils. Geotech Geol Eng 41:2325–2341. https://doi.org/10.1007/s10706-023-02400-0
Sun X, Li J (2023c) Tree root-soil interaction: field study of the effect of trees on soil moisture and ground movement in an urban environment. Plant Soil. https://doi.org/10.1007/s11104-023-06307-w
Sun X, Li J, Zhou AN (2017) Evaluation and comparison of methods for calculating Thornthwaite Moisture Index. Aust Geomech J 52(2):61–75
Sun X, Li J, Cameron DA, Zhou AN (2021) Field monitoring and assessment of the impact of a large eucalypt on soil desiccation. Acta Geotech. https://doi.org/10.1007/s11440-021-01308-4
Sun X, Li J, Cameron DA, Moore G (2022a) Sap flow monitoring of two Australian native tree species in a suburban setting: implications for tree selection and management. Plant Sci. https://doi.org/10.1016/j.plantsci.2022.111194
Sun X, Li J, Cameron DA, Moore G (2022b) On the use of sap flow measurements to assess the water requirements of three Australian native tree species. Agron J 12(1):52. https://doi.org/10.3390/agronomy12010052
Sun X, Li J, Cameron DA, Zhou AN, Bayetto P, Moore G (2023) Assessment of the influence of an Australian native eucalypt, Corymbia maculata, on soil desiccation in a temperate climate. Acta Geotech 18:5901–5919. https://doi.org/10.1007/s11440-023-02075-0
Swanson RH (1983) Numerical and experimental analyses of implanted-probe heat-pulse velocity theory. University of Alberta, Edmonton, Canada
Thayer RL, Maeda BT (1985) Measuring street tree impact on solar performance: a five-climate computer modelling study. Arboric J 11:1–12
Thorpe MR (1978) Net radiation and transpiration of apple trees in rows. Agric for Meteorol 19:41–57
Vandegehuchte MW, Steppe K (2013) Sap-flux density measurement methods: working principles and applicability. Funct Plant Biol 40:213–223. https://doi.org/10.1071/FP12233
Werk KS, Oren R, Schulze ED, Zimmermann R, Meyer J (1988) Performance of two Picea abies (L.) Karst. stands at different stages of decline. III Canopy Transpiration of Green Trees. Oecologia 76:519–524
Acknowledgements
This work was supported by the Australian Research Council via ARC Linkage Grant No. LP16160100649. The authors would like to thank the financial and technical support of the City of Knox, and FMG Engineering is also gratefully acknowledged.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. Jie Li was financially supported by Australian Research Council, LP16160100649.
Author information
Authors and Affiliations
Contributions
X.S. conceived the ideas and led the writing of the manuscript; J.L. selected the tree species and study sites; X.S. designed the methodology; X.S. collected and analyzed the sap flow data; all authors contributed to the interpretation of results. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by G. Locosselli .
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sun, X., Li, J. Patterns of water use by the Australian native Melaleuca styphelioides in urban environments and comparison of transpiration prediction by three different micrometeorological models. Trees 38, 493–506 (2024). https://doi.org/10.1007/s00468-024-02496-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-024-02496-8