
Abstract
In the cities, remnants of the vegetation that originally occupied these areas can be found. Exotic species have established in these relicts, and therefore these spaces require management aimed at recovering native vegetation. In this study, we evaluated the impact of the removing exotic species on the floristic composition of the remnants of an oak forest. We compared the abundance/cover and composition of exotic plants in a forest that has been managed through citizen participation with that of reference forests (both with remnants, but the former without protection and the latter with protection). We established 48 sampling sites to characterize the vegetation in three strata (upper, middle, and low) and monitored changes in composition in managed forest sites for a period of 3 years. The results indicate that the managed forest hosts different species than those found in the reference forests, with lower abundance and cover of exotic plants in the upper and lower strata. Regarding temporal changes, we observed shifts in floristic composition before and after management activities, and a reduction in the cover of exotic species before and after management, with consistently low cover three years later. This underscores the effectiveness of citizen-led management activities, offering a viable alternative for restoring native vegetation in urban environments.
Similar content being viewed by others


Introduction
Cities, which are conceived as socio-ecosystems, have gained great relevance for biodiversity due to the negative effects derived from the concentration of the human population, the production of pollutants, the change in land use and their exponential growth (Müller et al. 2013). The expansion of urban areas is usually associated with the destruction of the ecosystems that surround them. Despite this, some fragments of the original vegetation that occupied these spaces can remain within the cities and are called relict forests (Florgård 2007; Klaus and Kiehl 2021).
These spaces are of great importance for the people inhabiting cities, providing various ecosystem services and serving as museums of the ecosystems that once occupied those areas. Additionally, being part of the green infrastructure (Florgård 2007), they act as catalysts for socialization processes, such as social integration and interaction (Enssle and Kabisch 2020; Standish et al. 2013). For these reasons, conservation and, where appropriate, restoration must be priority activities. In cities, participatory restoration has proven to be an effective tool to help recover the remnants of vegetation, while helping people understand the importance of these relics for urban areas (Dolan et al. 2015; Krasny et al. 2015; Standish et al. 2013).
One of the most important activities in the recovery of relict vegetation is removing populations of invasive exotic species (Dolan et al. 2015). Some studies show that in urban environments, remove populations of exotic plants can be complicated since the dispersion of invasive exotic species to these areas is constant (Dolan et al. 2015; Gaertner et al. 2017). However, ignoring this activity can make the recovery process of urban green areas impossible or delayed, since the presence of exotic species prohibits the establishment of native species (Nguyen et al. 2021; Standish et al. 2013). The management of exotic species could be conducted through citizen participation, involving neighbors in removal work. Experiences of citizen participation in the removal of exotic species in urban areas have been studied (Connolly et al. 2013; Dolan et al. 2015; Heneghan et al. 2009), but little has been documented on changes in diversity after management by citizens over the years.
In Mexico City, numerous remnants of original vegetation exist (Ayala-Azcarraga et al. 2023). Among these, the most extensively researched ones, owing to their social, ecological and historical significance, are the vegetation fragments that have developed in the Xitle volcanic field, known as Pedregal de San Ángel (PSN) (Castillo-Argüero et al. 2004; Rzedowski 1994). While the vegetation in this region has been thoroughly investigated, there are limited studies on the oak forests that still persist within the urban area. In these remnants, issues have been reported due to the invasion of exotic species such as Apium graveolens, Eucalyptus spp., Hedera helix, Leonotis nepetifolia, Pennisetum clandestinum, and Vinca minor, to name a few (Castillo-Argüero et al. 2004; González-Hidalgo et al. 2001; SEDEMA 2011, 2018).
The remnants of the oak forest located in the southern area of Mexico City (CDMX) represent the last relicts within the urban area, which is why they are considered relevant ecosystems for the conservation of biodiversity. And although there are some studies of the oak forests in the southern peri urban area of the city (see for example: Bonfil Sanders et al. 2000), since 1953, no comprehensive studies have been carried out to characterize the diversity of these plant communities or to determine their degree of conservation within the urban area (Rzedowski 1994). Furthermore, it is unknown if management activities for the oak forest are effective for minimizing exotic species abundance and promoting native biodiversity.
The aim of this study is to evaluate the effect of management activities carried out by citizens on the floristic composition of plant communities in an oak forest compared to sites without intervention. To do this: (i) we compare the composition and abundance of exotic species between the managed forest and the reference forests (found in nearby protected natural areas) and (ii) we determine if there are changes in composition and dominance of exotic species in the managed forest over a period of 3 years.
Methods
The study area was limited to the relicts of a type of oak forest dominated by Quercus deserticola (OFQd) that currently exist in small areas in the south of CDMX, at altitudes ranging from 2350 to 2700 m a.s.l. The trees of this OFQd reaching a height of 6–10 m and grow in dense masses. This oak forest developed in clearings that were not covered by lava, which was the product of a volcanic eruption that occurred more than 1,700 years ago. In this lava field, other types of vegetation developed: towards the lower areas a xeric scrub, in the middle area different types of oak and mixed forests and towards the higher areas coniferous forests (Rzedowski 1994).
Within the urban area, there are relics of OFQd with different protection statuses according to local regulations. One is the Cerro de Zacatepetl Area of Environmental Value (CZ), and the other is the Bosque de Tlalpan Ecological and Cultural Zone Protected Natural Area (BT). These areas had kept protection against changes in land use and the regulation of human activities (such as species extraction, logging, recreation, among others) for at least the last 50 years, predating the urbanization of the area. Consequently, these areas are well-preserved (SEDEMA 2011, 2018). Outside of CZ and BT, there is another fragment of the OFQd in a 5 ha park with no formal protection called Viaje a la Naturaleza forest (VN). In VN, issues such as garbage problems, annual removal of all shrubby individuals, introduction of exotic species by some neighbors, and the presence of invasive exotic species were observed (Urroz 2022). Since 2020, various management activities have been carried out through citizen participation in this park (VN), as described below. Both CZ and BT, due to their well-preserved state, were considered reference forests, while VN, due to the activities undertaken since 2020, was labeled as a managed forest.
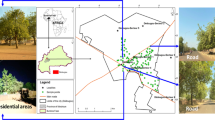
To characterize the plant communities, we stablished a total of 48 random sampling sites, of which, 20 sites were placed in VN in which management activities were carried out through citizen participation (managed forest). The other 28 sampling sites were established in the reference forests: 11 in CZ and 17 in BT (Fig. 1).

Map showing the cover of oak forests in 1953 according to (Rzedowski 1994) and the location of the sampling sites. (A) Sampling sites of the BT and CZ reference forests. (B) Close-up of the location of the sites in the managed forest (VN)
In each of the sampling sites, we stablished plots to characterize three plant strata. Each plot was made up of three squares: one of 30 × 30 m, one of 5 × 5 m and one of 1 × 1 m in which the vegetation of three vertical strata was recorded: upper, middle and low respectively. In the plot of 30 × 30 m (900m2), we recorded all the individual with normal diameter (DN: 1.3 m) ≥ 4 cm, in the plot of 5 × 5 (25 m2) we recorded the individuals with DN < 4 cm, and in the plot of 1 × 1 m (1 m2), individuals with a height < 1.3 m were recorded. We determined the species occurrence and abundance (in the plots of 1m2 the cover was determined) in all samples. In the 900 and 25 m2 squares the DN was recorded.
Management activities through citizen participation
In December 2020, the inhabitants of the surrounding area of the VN forest decided to implement management measures, with the aim of recovering the OFQd. The measures applied consist of the removal of dominant exotic species from the lower stratum (species names-acronyms used in figures: Hedera helix- He he, Jacaranda mimosifolia- Ja mi, Leonotis nepetifolia- Le ne, Pennisetum clandestinum- Pe cl y Vinca minor- Vi mi), in 2020 and 2021. Starting in 2022, all exotic species observed in the lower stratum began to be removed (Supplementary information). Additionally, the following recovery measures were implemented: the removal of impermeable surfaces, the delimitation of trails, the restriction of the mobility of users to the trails, the transplantation of native plant species in areas without tree cover, the extraction of garbage from the forest and the prohibition of throwing waste in the forest (Urroz 2022).
To remove exotic species, workshops on exotic plant identification were conducted for neighbors and workers. Two workshops were held before commencing the removal work, and six were conducted in 2021, with an attendance ranging from 10 to 30 people. The removal was carried out in 2021 through monthly or bimonthly work sessions and starting in 2022, the removal of an individual or population that was detected sporadically was carried out.
Evaluation of changes in vegetation composition (between forests and between seasons)
To evaluate the degree of conservation of the managed forest with respect to the reference forests, we visited all sampling sites in August 2023 and the vegetation in the three strata described above was recorded. To evaluate changes in composition over time (in different seasons), we visited the sampling sites within the managed forest in November 2020 (before management activities), August 2021, November 2021 and August 2023 (during management activities). In these visits, only the vegetation of the lower stratum was recorded since the removal activities of exotic species were limited to the plants of this stratum.
Statistical analysis
To compare the composition between the managed forest and the reference forests, we used a permutation analysis (PERMANOVA). Before performing the analysis, the abundance data were transformed using the Hellinger method (Borcard et al. 2018). To visualize these results, we performed a multivariate NMDS clustering analysis using a distance matrix calculated with the Bray-Curtis dissimilarity coefficient for each stratum. The PERMANOVA was performed using the adonis2 function from the Vegan package (Oksanen et al. 2013), for the NMDS the metaMDS function from the Vegan package was used (Oksanen et al. 2013). To compare the dominance of exotic species, we performed a Kruskal’s non-parametric test using abundance data (middle and upper stratum) or cover (low stratum). The kruskal.test function from the Vegan package was used (Oksanen et al. 2013). Subsequently, we performed an analysis to detect differences between the groups using the Wilcoxon test. The function pairwise.wilcox.test from the Vegan package was used (Oksanen et al. 2013).
Due to the marked seasonality of these forests, to analyze changes in composition before and after citizen intervention, comparisons were made between November 2020 and November 2021 (dry season) and between August 2021 and July-August 2023 (rainy season). Because the data were not normally distributed, a nonparametric Friedman test was performed for comparisons. We used the friedman.test function from the Vegan package to perform the test (Oksanen et al. 2013). To compare the composition, we performed a permutation analyzes (PERMANOVA) and an NMDS to visualize the similarity. For these last two analyzes we used the procedure and packages described above.
Results
We recorded a total of 155 species in the studied sites. In the Bosque de Tlalpan (BT), 78 species were found, of which 12 were exotic, in the Cerro de Zacatepetl (CZ) 65 species of which 9 were exotic, and in Viaje a la Naturaleza (VN) 78 species with 16 exotic (Supplementary information).
Differences between managed forest and the reference forests
We observed significant statistical differences in the abundance of individuals of exotic species between the sites of the managed forest (VN) and the reference forests (CZ, BT) for the upper (Kruskal Wallis x2 = 15.11, p = < 0.001) and low stratum (Kruskal Wallis x2 = 12.62, p = 0.002), but not for the middle stratum (Kruskal Wallis x2 = 3.80, p = 0.15). The Wilcoxon post-hoc tests indicate that in the upper stratum the abundance of exotic species was greater in the Bosque de Tlalpan and in the Cerro de Zacatepetl than in Viaje a la Naturaleza (BT-CZ p = 0.93; BT-VN p = 0.002; CZ-VN p = 0.01). In the lower stratum, a greater abundance of exotic species was observed in the Bosque de Tlalpan than in Cerro de Zacatepetl and Viaje a la Naturaleza (BT-CZ p = 0.05; BT-VN p = 0.002; CZ-VN p = 1) (Fig. 2).

Box and whisker plot of the abundance or cover of exotic species in each of the forests studied, (a) upper stratum (plot area 900 m2), (b) middle stratum (plot area 25 m2), (c) low stratum (plot area 1 m2). Capital letters (A, B) indicate significant statistical differences between sites
The species composition was different between the sites in all the strata evaluated (upper stratum: pseudo F = 12.30, p = < 0.001; middle stratum: pseudo F = 4.58, p = < 0.001; low stratum: pseudo F = 4.83, p = < 0.001) (Fig. 3).

Graph of the non-metric dimensional scaling analysis of the plant communities in the 3 forests for the (a) upper stratum, (b) middle stratum, and (c) low stratum. The location of the species in two-dimensional space is indicated with an asterisk, exotic ones in black and native ones in gray. Only native species that were observed at more than 3 sites and all exotic species are shown (abbreviations of names are found in Supplementary information). The bigger circles are the centroids
Temporal changes (Pre and post citizen intervention)
We observed differences in species composition before and after management, irrespective of the sampling season. We find that the cover of exotic species was lower in the second dry season (one year after management) than in the first (before management) (Friedman x2 = 8.89, p = < 0.01). We did not observe differences were observed between the first (one year after management) and the second (two years after management) rainy season (Friedman x2 = 0.08, p = 0.78) (Fig. 4).

Changes in the cover of exotic species in the lower stratum for different seasons in the forest with management (VN). The letters indicate significant statistical differences between the seasons (A, B, C)
The species composition was different between the first (before management) and second (one year after management) dry season (pseudo F = 3.70, p = < 0.001) and for the first (one year after management) and second (two years after handling) rainy season (pseudo F = 7.18, p = < 0.001) (Fig. 5).

Graph of the non-metric dimensional scaling analysis of the lower stratum communities before management (Nov 2020) and after management (Nov 2021, Aug 2021, Aug 2023). (a) dry season, (b) rainy season. The location of the species in two-dimensional space is indicated with an asterisk, exotic ones in black and native ones in gray. Only native species that were observed at more than 3 sites and all exotic species are shown (abbreviations of names are found in Supplementary information). The bigger circles are the centroids
Discussion
Citizen participation in the maintenance of areas with relict vegetation within urban areas is essential for the conservation of biodiversity in cities (Klaus and Kiehl 2021; Krasny et al. 2015; Standish et al. 2013). This study shows that activities such as the management of exotic species are important in the recovery/restoration tasks of these spaces. In previous works, it has been found that recovery activities such as tree transplantation, propagule collection (Krasny et al. 2015), monitoring (Callaghan et al. 2019) and the identification of exotic species for management (Jordan et al. 2012) can be carried out successfully by citizens.
This study focuses on documenting that the removal of exotic species by citizens in urban green areas is successful over time and that this management generates changes in the composition of native species. Works such as that of Dolan et al. (2015), have evaluated success for short periods of time and have found that the management of exotic species by residents of a green urban area in Indiana, United States, helped control an invasive exotic species (Lonicera maackii). The data obtained in our study demonstrate that in the managed forest sites (VN) the average cover of exotic species in the lower stratum was less than or equal to that of the reference forest sites (BT and CZ). Furthermore, the cover of exotic species decreased before and after management activities carried out by citizen, changing from 33.3% before the implementation of management activities to 7.5% after two and a half years of management. An example of the decrease in the cover of invasive exotic species over time that we quantified is in the case of Pennisetum clandestinum, which was more abundant before the management activities, as shown in the ordering analyses.
It is important to note that, despite the observed reduction in cover during the analysis period, management activities did not eliminate the presence of exotic species in the managed forest. This could be attributed to the dispersion of exotic species into the managed forest after the commencement of the work. This phenomenon was observed in the case of Anagyris arvensis and Acalypha indica, both exotic species recorded in 2023 but not in 2020 or 2021. The dispersal of exotic species towards urban green areas is a common process in cities (Dolan et al. 2015; Müller et al. 2013). In the area, the arrival of exotic species has been documented in green areas with no historical records of their presence 70 years ago (Castillo-Argüero et al. 2004). Additionally, the lower layer is highly susceptible to being occupied by exotic species. A study in urban forests on the northern Atlantic coast of the United States found that over time, different species could colonize these areas, and the lower stratum was the most vulnerable to the invasion of exotic plants (67% of recorded individuals were exotic species) (Bierzychudek 2020), both A. arvensis and A. indica belong to this stratum. This suggests that although management activities have effectively reduced the coverage of exotic species, the continuity of this trend in the long term is uncertain if management activities cease (Vidra et al. 2007).
Another relevant point to consider is the challenge of detecting inconspicuous or low-abundant exotic species by citizens involved in management activities. This was evident in the case of Chenopodium murale, Geranium seemannii, Trifolium repens, and Stellaria media, which were recorded in nearly all samplings since 2021. These species were not highly abundant or frequent in the samplings and typically do not exceed 20 cm in height (except for C. murale, which reaches 60 cm) (de Rzedowski and Rzedowski 2001). The difficulty in identifying inconspicuous species by citizens has been noted in previous studies, and it is likely that with greater citizen specialization in the detection of exotic species, rare or very small species can be accurately identified (Jordan et al. 2012).
The fact that complete elimination of exotic species from the managed forest has not been achievable, either due to propagule influx from other sites or the challenge of detecting inconspicuous exotic species, does not undermine the success of management activities. Similar reductions have been documented in management efforts by government authorities and scientific experiments, as evidenced by studies such as Bierzychudek 2020; Johnson and Handel 2016 and Vidra et al. 2007. For instance, a restoration project conducted in urban forests in New York reported a 50% coverage of three exotic plant species in the lower stratum. After 20 years of management, their coverage reduced to 7% (Johnson and Handel 2016). In our study, we observed that exotic species had a coverage of 33.3% before management, eight months after management (Aug-2021), it decreased to 10.4%, and two and a half years later (Aug-2023), it further diminished to 7.5%.
Regarding the contrasts in the composition between the managed forest (VN) and the reference forests (BT and CZ), the results show that the similarity in the composition of the upper and low stratum of some sites in the reference forests was higher with the managed forest sites than with the reference forest sites themselves. This result is given by two phenomena: (i) the greater similarity of some sites in the reference forests due to the presence of exotic species that were used in reforestations carried out in the middle of the last century (such as Eucalyptus spp. and Acacia spp.) (Rzedowski 1994). Furthermore, in the sites with reforestation, the dominance in the lower stratum of an invasive exotic grass species, P. clandestinum, was observed, probably favored by the greater opening of the canopy that tree individuals of Eucalyptus spp. have. (Livesley et al. 2014) and by the allelopathic effect that these species have in the native plants of this stratum (Castillo-Argüero et al. 2004). And (ii) the greater similarity between some sites in the reference forests with the managed forest sites that were dominated by native species, such as Quercus spp. and Eysenhardtia polystachya in the upper stratum, and Cissus sicyoides, Dicliptera peduncularis, Dioscorea galeottiana, Gaudichaudia cynanchoides and Salvia mexicana in the lower stratum. All of these species had been recorded before the area was urbanized (Rzedowski 1994). All of the above indicates that the composition of the managed forest is similar to the sites with the highest degree of conservation of the reference forests.
The differences in composition between the different seasons of the managed forest indicated that the floristic composition changes both in the dry season and in the rainy season. These differences may be related to the decrease in the cover of exotic species such as P. clandestinum and Hedera helix, which were more abundant in the first seasons of both the dry (November 2020) and the rainy (August 2021). Additionally, ordination analyzes indicate that these differences are also related to changes in the cover of native species such as Brachypodium mexicanum, C. cisyoides and G. cynanchoides. These species had low cover before the management activities (November 2020), but for the subsequent sampling seasons (August 2021, November 2021 and August 2023) their cover increased significantly. The above indicates that in the managed forest not only did the cover of exotic species decrease, but also the cover of some native species increased.
Conclusion
Our findings demonstrate that citizen-led management significantly reduces the coverage of exotic species, and if sustained, this reduction persists over time. The management was effective for the most abundant exotic species, while for the less abundant or inconspicuous ones, it did not show the same effectiveness. Therefore, if the objective is to eliminate all exotic species, specialized support will be required, both in knowledge and resources. Based on this, it is recommended that citizens participate in the recovery efforts of relict forests and urban green areas, not only through the removal of exotic species. It is likely that citizens can support the propagation of native species, engage in soil improvement activities (e.g., generating compost), monitor pests, contribute to reforestation, and participate in environmental education activities. These activities should be planned through workshops and in collaboration with academic institutions and the government.
References
Ayala-Azcarraga C, Diaz D, Fernandez T, Cordova-Tapia F, Zambrano L (2023) Uneven distribution of urban green spaces in relation to marginalization in Mexico City. In Sustainability (Vol. 15, Issue 16). https://doi.org/10.3390/su151612652
Bierzychudek P (2020) Passive recovery of an urban forest in the Pacific Northwest after removal of invasive plants. Urban Ecosyst 23(5):1023–1038. https://doi.org/10.1007/s11252-020-00966-8
Bonfil Sanders C, Rodríguez Vega H, Peña Ramírez V (2000) Evaluación Del efecto de plantas nodrizas en El establecimiento de una plantación de Quercus L. Revista Mexicana De Ciencias Forestales 25(88 SE-Artículo Científico):59–73. https://cienciasforestales.inifap.gob.mx/index.php/forestales/article/view/923
Borcard D, Gillet F, Legendre P, Borcard D, Gillet F, Legendre P (2018) Association measures and matrices. Numer Ecol R 35–57
Callaghan CT, Major RE, Lyons MB, Martin JM, Wilshire JH, Kingsford RT, Cornwell WK (2019) Using citizen science data to define and track restoration targets in urban areas. J Appl Ecol 56(8):1998–2006. https://doi.org/10.1111/1365-2664.13421
Castillo-Argüero S, Montes-Cartas G, Romero-Romero MA, Martínez-Orea Y, Guadarrama-Chávez P, Sánchez-Gallén I, Núñez-Castillo O (2004) Dynamics and conservation of the flora of the desert scrub of the Ecological Reserve of the Pedregal de San Angel (D. F., Mexico). Bot Sci 0(74 SE-ARTICLES / ARTÍCULOS). https://doi.org/10.17129/botsci.1686
Connolly JJ, Svendsen ES, Fisher DR, Campbell LK (2013) Organizing urban ecosystem services through environmental stewardship governance in New York City. Landsc Urban Plann 109(1):76–84. https://doi.org/10.1016/j.landurbplan.2012.07.001
Dolan RW, Harris KA, Adler M (2015) Community involvement to address a long-standing invasive species problem: aspects of Civic Ecology in Practice. Ecol Restor 33(3): 316 LP – 325. https://doi.org/10.3368/er.33.3.316
Enssle F, Kabisch N (2020) Urban green spaces for the social interaction, health and well-being of older people— an integrated view of urban ecosystem services and socio-environmental justice. Environ Sci Policy 109:36–44. https://doi.org/10.1016/j.envsci.2020.04.008
Florgård C (2007) Preserved and remnant natural vegetation in cities: a geographically divided field of research. Landsc Res 32(1):79–94. https://doi.org/10.1080/01426390601097750
Gaertner M, Wilson JRU, Cadotte MW, MacIvor JS, Zenni RD, Richardson DM (2017) Non-native species in urban environments: patterns, processes, impacts and challenges. Biol Invasions 19(12):3461–3469. https://doi.org/10.1007/s10530-017-1598-7
González-Hidalgo B, Orozco-Segovia A, Diego-Pérez N (2001) The vegetation of the Lomas Del Seminario Ecological Reserve, Ajusco, México. Bot Sci 0(69). https://doi.org/10.17129/botsci.1648
Heneghan L, Umek L, Bernau B, Grady K, Iatropulos J, Jabon D, Workman M (2009) Ecological research can augment restoration practice in urban areas degraded by invasive species—examples from Chicago wilderness. Urban Ecosyst 12(1):63–77. https://doi.org/10.1007/s11252-008-0057-8
Johnson LR, Handel SN (2016) Restoration treatments in urban park forests drive long-term changes in vegetation trajectories. Ecol Appl 26(3):940–956. https://doi.org/10.1890/14-2063
Jordan RC, Brooks WR, Howe DV, Ehrenfeld JG (2012) Evaluating the performance of volunteers in mapping invasive plants in public conservation lands. Environ Manage 49(2):425–434. https://doi.org/10.1007/s00267-011-9789-y
Klaus VH, Kiehl K (2021) A conceptual framework for urban ecological restoration and rehabilitation. Basic Appl Ecol 52:82–94. https://doi.org/10.1016/j.baae.2021.02.010
Krasny ME, Silva P, Barr C, Golshani Z, Lee E, Ligas R, Mosher E, Reynosa A (2015) Civic ecology practices: insights from practice theory. Ecol Soc 20(2). http://www.jstor.org/stable/26270185
Livesley SJ, Baudinette B, Glover D (2014) Rainfall interception and stem flow by eucalypt street trees – the impacts of canopy density and bark type. Urban Forestry Urban Green 13(1):192–197. https://doi.org/10.1016/j.ufug.2013.09.001
Müller N, Ignatieva M, Nilon CH, Werner P, Zipperer WC (2013) Patterns and trends in urban biodiversity and landscape design BT - urbanization, biodiversity and ecosystem services: challenges and opportunities: a global assessment (T. Elmqvist, M. Fragkias, J. Goodness, B. Güneralp, P. J. Marcotullio, R. I. McDonald, S. Parnell, M. Schewenius, M. Sendstad, K. C. Seto, & C. Wilkinson (eds.); pp. 123–174). Springer Netherlands. https://doi.org/10.1007/978-94-007-7088-1_10
Nguyen N-A, Eskelson BNI, Gergel SE, Murray T (2021) The occurrence of invasive plant species differed significantly across three urban greenspace types of Metro Vancouver, Canada. Urban Forestry Urban Green 59:126999. https://doi.org/10.1016/j.ufug.2021.126999
Oksanen J, Simspson GL, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’Hara R, Solymos P, Stevens MHH, Szoecs E, Wagner H, Barbour M, Bedward M, Bolker B, Borcard D, Carvalho G, Chirico M, Caceres M, De, Durand S, Weedon J (2013) Package ‘Vegan’. Community Ecology Package. https://cran.r-project.org/package=vegan
Rzedowski J (1994) Vegetación Del Pedregal de San Ángel. In: Rojo A (ed) Reserva Ecológica El Pedregal de San Ángel: Ecología E Historia natural y manejo. Universidad Nacional Autónoma de México, pp 9–65
Rzedowski GC, Rzedowski J (2001) Flora fanerogámica del Valle de México. Comisión Nacional Para El Estudio de La Biodiversidad e Instituto de Ecología, AC. México, 983
SEDEMA (2011) Programa de manejo del área natural protegida Zona Ecológica y Cultural Bosque de Tlalpan
SEDEMA (2018) Programa de manejo del área de valor ambiental Bosque Urbano Cerro Zacatepetl (p. 50)
Standish RJ, Hobbs RJ, Miller JR (2013) Improving city life: options for ecological restoration in urban landscapes and how these might influence interactions between people and nature. Landscape Ecol 28(6):1213–1221. https://doi.org/10.1007/s10980-012-9752-1
Urroz R (2022) Habitar Un ecosistema: la experiencia comunitaria en torno al bosque de jardines en la Montaña De La Ciudad De México. Investigaciones Geográficas 109. https://doi.org/10.14350/rig.60670
Vidra RL, Shear TH, Stucky JM (2007) Effects of vegetation removal on native understory recovery in an exotic-rich urban forest. J Torrey Bot Soc 134(3):410–419. https://doi.org/10.3159/1095-5674(2007)134[410:EOVRON]2.0.CO;2
Acknowledgements
To Rubén Salinas-Galicia, Mónica M. Vázquez-Medrano, Sofía Vizcaino for their support in the field work. To Herminia Torres, Mónica Ornelas, Francisco Moya, Raúl Reyes, Úrsula Huerta and the members of the Comité de Ecología de Jardines en la Montaña for carrying out the management activities. To the Asociación de Jardines en la Montaña, to the Direction of the ANP ZEC Bosque de Tlalpan and the ANP Cerro de Zacatepetl for allowing me access to the forest.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by F-G N., G, E. and T I. The first draft of the manuscript was written by F-G N. and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Flores-Galicia, N., Gutiérrez, E. & Trejo, I. Effect of exotic species management on the recovery of relict forests through citizen participation. Urban Ecosyst (2024). https://doi.org/10.1007/s11252-024-01535-z
Accepted:
Published:
DOI: https://doi.org/10.1007/s11252-024-01535-z