
Abstract
Research evaluating the impact of silvopastoral systems on physical and biological properties of Amazonian soils is scarce. Thus, this study aimed to compare the soil carbon storage potential and physical and chemical soil properties of silvopastoral systems (SPS) and conventional pastoral systems (CPS) in the San Martin region of Peru. Using the Walkley and Black method, we analyzed soil organic matter at two different depths (0–15 cm and 15–30 cm). In addition, bulk density, soil moisture, total porosity, and mechanical resistance were measured in both systems. The highest (P < 0.05) carbon stocks were reported at 0–15 cm of depth with values of 31.4 Mg ha−1 and 34.4 Mg ha−1 for CPS and SPS, respectively. At 15–30 cm depth, the total carbon stock was higher for SPS, with 29.12 Mg ha−1, than for CPS, which had 26.4 Mg ha−1. Despite the absence of statistically significant differences, soil carbon stocks were higher in SPS. No significant differences in soil moisture were found between systems, although soil moisture was slightly greater in SPS than CPS (28 and 25%, respectively). The CPS had 59% of the total porosity, which was higher than the SPS. Mechanical resistance was lower in SPS (2.15 kg/cm2) than in CPS (2.33 kg/cm2) at 10 cm of depth. These results indicated that the SPS has the potential to store more carbon and improve physical and chemical traits in the soil than the CPS.
Similar content being viewed by others
Introduction
The growing human population and subsequent demand for agricultural products has accelerated land use change and degradation (Borrelli et al. 2017; Winkler et al. 2021). It is estimated that at least 70 Mkm2, or > 50% of the Earth’s ice-free land area (Hooke and Martın-Duque JF 2012), have been directly modified by humans. Degraded lands, or those in which deteriorating soil properties impact function, comprise approximately 24% of the global land area (35 Mkm2, Smith et al. 2016).
In the Peruvian Amazon, there are 9.6 million deforested hectares, of which only 1.9 million are used for agriculture and livestock production. The rest is underutilized, abandoned, and degraded with depleted soil carbon stocks. In addition, lands in agricultural production produce one-third to one-half of what they could produce if they were well managed (Dourojeanni 2022).
The IV National Agricultural Census in Peru documented 358,270.00 ha in pasture for livestock production (INEI 2012). Overstocking and overgrazing are common challenges in these areas. In addition, Amazonian cattle ranchers lack access to appropriate cattle breeds, technologies, and pasture management plans (Basilio and Malpartida 2022). All this causes the pastures to deteriorate rapidly, accelerating soil degradation processes (Alegre et al. 2019; Dourojeanni 2022).
Environmental, geological, and biotic factors such as the composition and density of plant and animal species (Fernández et al. 2013) have a direct influence on soil carbon reserves (Vesterdal et al. 2013). Anthropogenic activities have a significant effect on the rate of change in soil organic carbon (SOC) stock (Don et al. 2011), and land use conversion is a significant factor in the change of SOC stock and global CO2 emissions (IPCC 2014). Del Galdo et al. (2003) demonstrated that soils from annual agricultural croplands had 48% lower SOC content in the 10 cm soil layer compared to soils on lands that had been converted from annual cropland to permanent grassland or following afforestation.
Silvopastoral systems (SPS) are an alternative land use practice that integrates agricultural production with provisioning of ecosystem services, including carbon sequestration (Contreras-Santos et al. 2020). These systems reduce soil degradation through the incorporation of organic matter from cattle manure and plant litter that accumulates on the soil surface, improving the soil as a carbon sink (Alegre et al. 2019). In addition, SPS improve the nutritive quality of the pasture, reducing enteric methane emissions from cattle and allowing natural regeneration in degraded areas (Ibrahim et al. 2007).
The San Martin Amazon region has 70,000 hectares of pasture, affirming the great potential of the region for pasture livestock production. However, this activity has generated a high rate of deforestation. Therefore, the implementation of SPS has been a production option under an integrated management system that is socially, ecologically, and economically sustainable (Roque-Alcarraz et al. 2022).
Aligned with appropriate management practices, the tree, shrub, and herbaceous component of SPS can result in increased contributions of biomass and soil cover, which promotes nutrient cycling, biological activity, C sequestration, and soil storage (Olaya-Montes et al. 2020). Increasing SOC fostered by plant biomass (above and below ground) can improve the physical properties and processes of the soil (Vázquez et al. 2020), thereby promoting the growth of plants that might otherwise be restricted by compaction caused by animal trampling (Batista et al. 2019).
When conventional pastoral systems (CPS), are converted to SPS, the physical characteristics of the soil may improve (Polanía-Hincapié et al. 2021). Soil bulk density decreases and organic matter increases due to the decomposition of the litter and the intense root turnover (Junior et al. 2014), subsequently minimizing soil compaction (Martinkoski et al. 2017).
In the Amazonian region, few studies have evaluated carbon stocks and soil properties in SPS. Therefore, this study had two objectives: (1) to determine the soil carbon stock potential, and physical and chemical soil properties of conventional pastoral systems (CPS) compared with SPS at two depths in the San Martin region; and (2) to evaluate the potential of SPS for generating ecosystem services, such as carbon sequestration, while sustaining agricultural production.
Materials and methods
Site and systems description
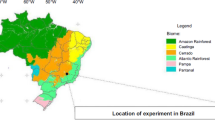
The experiment was conducted on 22 farms located in the district of Cuñumbuqui, in the region of San Martin, Peru (Fig. 1). This district is 200 m above sea level and has seasonal temperatures, relative humidity, and rainfall monthly (average of dry and wet rainy season) of 27.5 °C, 73.5%, 42 mm, respectively (Fig. 2). In addition, the district of Cuñumbuqui in the San Martín region is characterized by a tropical climate of rainy, semi-warm, and humid savannah, classified as a tropical dry forest ecosystem (Holdridge 1967). The data were collected in the dry season (July 2022).

Location map of soil collection points in Cuñumbuqui district. The orange dots next to the letters represent the collection points. The black area on the map represents the San Martin region of Peru, while the white dots within this area indicate the specific study zone. (Color figure online)

Source SENAMHI (2023). A Rainfall, B Temperature and relative humidity
Monthly climate conditions in the San Martin region during the collection year
Twenty-two farms (11 CPS and 11 SPS) were randomly selected from a pre-selection list of 72 farms from Cuñumbuqui that had been categorized following site visits. The criterion for defining CPS was the presence of pastures as a monoculture, with less than 15% tree cover, and SPS were pastures with tree and shrub cover exceeding 15% of the area devoted to cattle production.
Both livestock systems shared characteristics, including herd genotype, a cross of the Gyr breed with Holstein, Jersey, Brown Swiss, and Brahman, among others; the Brachiaria brizantha grass varieties Marandú, Xaraés, and Mulato were the primary forage source. The principal tree species identified in the SPS were Cecropia membranacea, Vochysia vismiifolia, Ceratonia siliqua, Manilkara bidentata, Juniperus thurifera, and Macrolobium acaciaefolium, with Gliricidia sepium and Erythrina poeppigiana representing the principal shrubs. A small number of dispersed trees, including Inga edulis, Guazuma ulmifolia, and Barbeyana cogniaux, were identified in the CPS.
Assessment of the soil physical and chemical properties
In both systems studied, the physical soil properties were analyzed. The soil analysis methods used are outlined in Table 1. The physical properties included bulk density (g/cm3), soil texture, moisture content (%), total porosity (%), and mechanical resistance (kg/cm2). Carbon stocks (t/ha) were determined by the Walkley and Black method. Soil organic matter fractionation was determined by extraction in alkaline medium and then separation in acid medium of labile sources (fulvic and humic acid) and recalcitrant (humins) (McLauchlan and Hobbie 2004). These parameters were analyzed at the Soil, Plant, and Water Analysis Laboratory of the La Molina National Agrarian University (UNALM) in Lima, Peru.
Source: Bazan (2017)
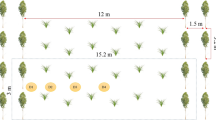
At each soil collection point (Fig. 2), four soil samples were extracted from two pits, two at 0–15 cm and the two others at 15–30 cm deep. Soil samples were collected randomly under the treetops, while in others areas were collected in open fields. Bulk density, mechanical resistance of the soil, and other chemical properties listed in Table 1 were analyzed at each sampling site for each depth. The soil degradation or enhancement status was evaluated based on the differences in C stocks.
Using a metal cylinder with a known volume of 100 cm3, two samples were obtained from each horizon of the soil profile to determine the bulk density (BD, g/cm3, Nachtergaele et al. 2023). Wet weights of soils were recorded before it has been dried in an oven for 48 h at 105 °C; after this procedure, dry weights were recorded. Finally, the BD was calculated by dividing the data for the dry soil mass by the cylinder’s volume.
The mechanical resistance (MR, kg/cm2) was measured using a manual pocket penetrometer at six intervals ranging from 5 to 30 cm deep of soil surface according methodology described by Dane and Top (2002). The soil moisture (H, %) of the same sample used for BD was assessed using the gravimetric approach (Porta et al. 2003). Soil carbon content was obtained by dividing the organic matter content by the humification factor of 1.724 (Walkley and Black 1934) and calculated with the bulk density and depth sampled and expressed in t C ha−1 for each of the two depths.
Additionally, the chemical fractionation of organic matter was done by sequentially separating humic substances based on how well they dissolved in acidic and alkaline media. This procedure allowed the quantification of the carbon in humic acids (CAH), fulvic acids, and non-humic substances (CNH + FAF), using the methodology described by Walkley and Black (1934).
Statistical design
A randomized complete block design with a factorial 2 × 2 arrangement was used. The SPS and CPS were replicated eleven times (each farm represented a replicate). The first factor was the livestock system (SPS and CPS), and the second was the sampling depth (0–15 cm and 15–30 cm). The data were analyzed using PROC MIXED from SAS (SAS for Windows V 9.4). Analysis of variance (ANOVA) was applied to each of the evaluated parameters. Means were compared using the SAS PDIFF procedure adjusted for Tukey’s test. Differences were declared significant at P ≤ 0.05.
Results
Soil texture
There was no significant effect for soil texture at 0–15 cm in SPS and CPS. From 15–30 cm there was a small but significant effect with sand and clay. At this depth, the soil had greater percent clay in the SPS than in the CPS, while the CPS had greater percent sand than SPS (Table 2).
Carbon stock in SPS and CPS
There were no significant differences between SPS and CPS carbon reserves; however, independent of the system, the greatest carbon accumulation was found at 0–15 cm depth with values of 32.9 Mg C ha−1. The average carbon content of the SPS was 29.19 Mg C ha−1 compared to 26.40 t C ha−1 for the CPS (Table 2).
Soil moisture and total porosity
No interaction was found between soil depth × livestock system (Table 2). Even though no significant differences were found in moisture for each system (P = 0.4320) and depth (P = 0.3356), a numerical difference was evident, being greater in the SPS than in the CPS, at 0–15 and 15–30 cm depth (28 and 27%, respectively).
There was no interaction (P > 0.05) between total porosity and depth. However, total porosity was significantly different between systems, and also significantly different between the 0–15 cm and 15–30 cm depths. Table 2 demonstrates that 58% of the pores are located within the first 15 cm of soil surface, which is greater (P < 0.05) than the 56% estimated at 30 cm. The CPS had 59% of the total porosity, which was greater (P < 0.05) than the SPS.
Bulk density (BD)
Table 2 demonstrates that the soil bulk density was greater (P < 0.05) at 15–30 cm than at 0–15 cm depth. The data demonstrates that the SPS soils had higher BD levels (P < 0.05) than CPS soils.
Organic matter (OM)
There were significant differences observed between both systems within the 0–15 cm range. However, during the 15–30 cm range, the CPS system demonstrated greater concentration over the SPS system (Table 2).
Chemical fractionation of OM
There was an interaction (P < 0.05) between System × Depth for the humin percentages (Table 2). The CPS at 15–30 cm has the maximum humin concentration at 3.1%, surpassing the SPS at the same depth and at 0–15 cm. There were no significant differences between SPS and CPS for fulvic and humic acids. The highest humic acid content (0.77%) was found in the top 15 cm of soil, while fulvic acids were found at 15–30 cm depth (Table 2).
Mechanical resistance
There was an interaction between System × Depth to mechanical resistance as an indicator of soil compaction, shown in Fig. 3. The SPS exhibited a lower mechanical resistance at 10 and 15 cm than the CPS.

Soil mechanical resistance (kg/cm2) in the soil of two livestock systems. CPS = Conventional pastoral system, SPS = Silvopastoral system. *Indicate interaction (P < 0.05) between livestock system and depth at the 5% level of significance
Discussion
Agroforestry, including silvopasture, has been identified as an important land use strategy for increasing soil and vegetation carbon stocks and mitigating climate change (Chapman et al. 2020). Silvopastoral systems can affect soil properties, increasing carbon in soil organic matter (SOM) through litter decomposition following leaf deposition. This turns more stable the granulometric fractions (silt + clay) of soil organic matter, confirming the importance of the silt and clay fractions for the aggregation of SOM (Guardiola et al. 2017) and physical protection of soil C. This effect was observed in our study, where SPS improved the clay percentage in 15–30 cm depth. In addition, according to de Souza Almeida et al. (2021) the lower C retention in sand is influenced by soil texture and the greater protective effect of the C by silt + clay. The lowest C content associated with sand is found in SPS. Soils that store more carbon have a more clayey silt texture and this is usually related to the effect of SPS, which during its implementation increased the C content of the soil by 7% in relation to native vegetation (de Souza Almeida et al. 2021).
The contribution of SPS on soil carbon stock is confirmed as well by Contreras-Santos et al. (2020) who reported that SPS accumulate between 60.6 and 65.1 Mg ha−1 of organic carbon in the soil, compared to typical treeless grazing systems, which accumulate just 28.3 Mg ha−1 of C. Similarly, Rojas et al. (2009) obtained values of 121.7 and 121.2 Mg ha−1 of C for silvopastoral systems with Pithecellobium saman and Diphysa robinioides associated with B. brizantha, respectively, in comparison to 87.7 Mg ha−1 of C for B. brizantha monoculture pasture. In this study, although we did not observe a significant difference in the soil carbon stock between systems, there is evidence of a greater stock potential for the SPS. This may be attributed to the decomposition of tree leaves deposited in the soil, transforming over time into soil aggregates that are distributed in the fractions of soil organic matter.
A number of studies have documented a significant improvement in soil carbon stocks in SPS compared to CPS (Junior et al. 2014; Martinkoski et al. 2017; Alegre et al. 2019; Contreras-Santos et al. 2020; Polanía-Hincapié et al. 2021; Roque-Alcarraz et al. 2022). In our study, no significant differences were found in the carbon stock between systems, which may be due to the presence of trees on farms categorized as CPS. Although tree cover was restricted to less than 15% of the total area of the farm, this minimal presence of trees could have increased the levels of carbon in soil layers. This is confirmed by Morales-Ruiz et al. (2021) who mentions that scattered trees and SPS with defined tree arrangements store more aboveground biomass and soil organic carbon than open pasturelands.
On the other hand, carbon varies greatly in soil layers and this is associated with soil texture and the roots of herbaceous, shrubby and tree plants. This is confirmed by Rojas et al. (2009) who demonstrated that the depth considerably influenced the organic carbon content of the soil, with the first 20 cm in both silvopastoral and conventional systems having the highest organic carbon content. On average, 55% of the soil’s total carbon is in the top 20 cm, compared to 25% and 20% at depths of 20 to 40 cm and 40 to 60 cm, respectively. Vasquez et al. (2020) discovered that at a depth of 0 to 15 cm, it was substantially greater than at a depth of 15 to 30 cm; in the first depth (0–15 cm), averages ranged from 108.85 to 76.73 Mg ha−1 of C, while in the second depth (15–30 cm), averages ranged from 40.20 to 24.57 Mg ha−1 of C. According to Ibrahim et al. (2007), soil carbon varies with depth and depends on the type of soil, its organic matter content, and the decomposition rate. The results obtained in this essay concur with these findings.
The increased soil moisture in SPS reported in this study is likely attributed to the function of tree shade in preventing soil moisture loss (da Silva et al. 2021). The low moisture obtained in both systems may be due to the scant precipitation reported during the season of data collection (July, Fig. 1). Due to the high likelihood of senescent material on the soil’s surface, the moisture percentage was greater at the top 15 cm than at 30 cm. In addition, Bucheli et al. (2013) note that tree species composition influences soil moisture, and the SPS had a different tree composition than the CPS.
The SPS appeared to have more moisture than the CPS, so it was anticipated that these sites would have larger pores. However, the SPS exhibited smaller pores. This is likely due to the fact that pore spaces may be filled with water, making them inaccessible for ventilation and soil moisture retention. In addition, certain soils have a denser, more compact structure that restricts the quantity of available pore space Hao et al. (2007). This information correlates with the higher apparent density and reduced total porosity of the SPS studied.
In addition, according to Sanchez (2021), the low bulk density values of the soil are influenced by high moisture contents and senescent material, such as tree leaf litter and pasture roots that accumulate in the superficial layer. Furthermore, Donoso (1994), indicates that pores with excellent drainage, aeration, and water infiltration have low BD values. Similarly, our results concur with Nachtergaele et al. (2023), which recommends that soils have a BD between 1.1 and 1.2 g/cm3. Regardless of system type, the lowest BD was discovered at depths between 0 and 15 cm, possibly due to the porous space, well-aerated soils, and the accumulation of plant remains on the soil surface (Alegre et al. 2017).
According to Junet et al. (2013), soil organic C is present between 20 and 100 cm deep. The soil organic matter is a heterogeneous mixture of organic compounds of various origins. Humins is the most stable fraction of soil organic matter and is considered the most resistant to microbial attack, as it is less active than humic acids and has lost a portion of the reactive groups (carboxyls and phenols) responsible for ionic exchange and the formation of stable aggregates (Conti and Giuffre 2011 as cited by Rios et al. 2016). The low microbial biomass in CPS may not be sufficient for humins degradation and carbon utilization.
The fraction of organic matter that is the most stable contains humic acids. It is the fraction that has the highest ionic exchange capacity and plays a significant role in the formation of stable aggregates by acting as a binder for the particles, and it is typically found in the top 0 to 15 cm of the soil, as in this study. In contrast, fulvic acids have the lowest molecular weight and the highest solubility; in general, they contain a greater proportion of aliphatic compounds than humic acids and humins (Rios et al. 2016). These characteristics make them more susceptible to washing and loss via diffusion in the profile following rain events. Fulvic acids are carried from the most superficial layers to deeper layers, with the deepest values found at a depth of 15 to 30 cm on this study. While this investigation did not identify any disparities in fulvic acid between the two systems, another investigation conducted by Loss et al. (2014) observed that SPS had a positive impact on the proportions of both fulvic acid and humic acid. The effect can be linked to the accumulation of organic matter from fallen leaves and the release of organic compounds by grassroots into the soil, known as rhizodeposition. The deposition of organic material with a high carbon-to-nitrogen ratio promotes the process of humidification of soil organic matter (SOM) and subsequently leads to the accumulation of organic matter in protected compartments, as noted by da Silva et al. (2020) and Lima et al. (2008).
The first 10 to 15 cm of the SPS in this study exhibited less mechanical resistance, most likely due to the contribution of leaf litter, moisture, and porous spaces. Between 20 and 30 cm, the mechanical resistance of both systems is comparable. This is likely due to soil moisture. Although the BD of the CPS was lower than that of the SPS, compaction typically results from an increase in BD due to the pressure exerted by cattle, which can lead to degradation of soil structure by decreasing aggregate stability (Sanchez 2021).
The classification criterion utilized for the SPS and CPS was based on the proportion of tree cover inside the livestock area, with a threshold of greater than 15% for SPS and less than 15% for CPS. The limited statistical differences between the SPS and CPS sites may be attributable to the existence of trees inside the CPS, which can influence the levels of organic matter, bulk density (BD), organic carbon, and mechanical resistance through the shading impact exerted by the scattered trees. This outcome aligns with expectations for a SPS. This finding may indicate that even a limited presence of trees has the potential to modify the physical properties of soil, facilitating the soil carbon sequestration.
Conclusions
The carbon reserves in both the SPS and CPS livestock systems were found to be greatest at a soil depth of 0–15 cm.
The SPS soils (1.19 g/cm3) had lower bulk density than the CPS (1.06 g/cm3) but lower mechanical resistance in the upper soil layers (2.15 kg/cm2), possibly allowing for increased water storage.
The chemical fractions of organic matter have a strong relationship with soil layers and fractions such as humin are susceptible to the effects that livestock systems can provide.
The absence of notable distinctions of carbon stock and some physical and chemical soil traits among systems may be attributed to soil heterogeneity within sites and the influence of scattered trees in the CPS. However, the numerical values among systems indicated that the SPS has the potential to store more carbon and improve physical and chemical traits in the soil than the CPS.
Despite not finding significant differences between the CPS and SPS in soil carbon stocks, it was evident that both systems can have an influence on the physical parameters of the soil, such as organic matter and on the fractions of organic matter, bulk density, soil texture, and mechanical resistance. The present study demonstrated that SPS probably influences the percentage of clay in the soil and causes the soil to have lower mechanical resistance in the first layers of the soil.
References
Alegre J, Lao P, Silva C, Schrevens E (2017) Recovering degraded lands in the Peruvian Amazon by cover crops and sustainable agroforestry systems. Peruv J Agron 1:1–7. https://doi.org/10.21704/pja.v1i1.1005
Alegre JC, Sánchez Y, Pizarro DM, Gómez C (2019). Manejo de los suelos con sistemas silvopastoriles en las regiones de Amazonas y San Martín. Extension material of Universidad Nacional Agraria la Molina. Print: Lima, Peru, pp 1–23. https://www.researchgate.net/publication/335541762_Manejo_de_los_suelos_con_sistemas_silvopastoriles_en_las_regiones_de_Amazonas_y_San_Martin
Basilio LHM, Malpartida NS (2022) Caracterización de la ganadería bovina del distrito de Codo de Pozuzo Provincia de Puerto Inca Región Huánuco. Universidad Nacional Daniel Alcides Carrión, Tesis para obtener el grado de Ingeniero Zootecnista
Batista PH, de Almeida GL, de Lima RP, Pandorfi H, da Silva MV, Rolim MM (2019) Impact of short-term grazing on physical properties of Planosols in Northeastern Brazil. Geoderma Reg 19:234. https://doi.org/10.1016/j.geodrs.2019.e00234
Borrelli P, Robinson DA, Fleischer LR, Lugato E, Ballabio C, Alewell C, Meusburger K, Modungo S, Schutt B, Ferro V, Bagarello V, Van Oost K, Montanarella L, Panagos P (2017) An assessment of the global impact of 21st century land use change on soil erosion. Nat Commun 8:2013. https://doi.org/10.1038/s41467-017-02142-7
Bucheli P, Benjamin T, Rusch G, Ibrahim M, Casals P, Sánchez D, Pugnaire F (2013). Estrategias de los árboles para el uso eficiente del agua y tolerancia a la sequía en sistemas silvopastoriles. Agroforestería en las Américas, N°50. https://brage.nina.no/nina-xmlui/handle/11250/2561400
Chapman M, Walker WS, Cook-Patton SC, Ellis PW, Farina M, Griscom BW, Baccini A (2020) Large climate mitigation potential from adding trees to agricultural lands. Glob Change Biol 26:4357–4365. https://doi.org/10.1111/gcb.15121
Contreras-Santos JL, Martínez-Atencia J, Cadena-Torres J, Falla-Guzmán CK (2020) Evaluación del carbono acumulado en el suelo en sistemas silvopastoriles del Caribe colombiano. Agron Costarric 44:29–41. https://doi.org/10.15517/rac.v44i1.39999
Conti ME, Giuffre L (2011) Edafología, Bases y Aplicaciones Ambientales. Universidad de Buenos Aires, CABA, p 623
da Silva IAG, Dubeux JC, Santos MVF, de Mello ACL, Cunha MV, Apolinário VX, de Freitas EV (2021) Tree canopy management affects dynamics of herbaceous vegetation and soil moisture in silvopasture systems using arboreal legumes. Agronomy 11:1509. https://doi.org/10.3390/agronomy11081509
da Silva JRM, Ensinas SC, Barbosa GF, Rezende JVO, Barreta PGV, Zuffo AM (2020) Total organic carbon and the humic fractions of the soil organic matter in silvopastoral system. Rev Bras De Ciênc Agrár 15:1–6. https://doi.org/10.5039/agraria.v15i2a6874
de Souza Almeida LL, Frazão LA, Lessa TAM, Fernandes LA, de Carvalho Veloso ÁL, Lana AMQ, Ferreira EA (2021) Soil carbon and nitrogen stocks and the quality of soil organic matter under silvopastoral systems in the Brazilian Cerrado. Soil Tillage Res 205:104785
Dane JH, Top CG (eds) (2002) Methods of soil analysis, Part 4 physical methods, vol 20. Wiley, New York
Del Galdo I, Six J, Peressotti A, Francesca CM (2003) Assessing the impact of land-use change on soil C sequestration in agricultural soils by means of organic matter fractionation and stable C isotopes. Glob Change Biol 9:1204–1213. https://doi.org/10.1046/j.1365-2486.2003.00657.x
Donoso ZC (1994). Ecología forestal. El bosque y su medio ambiente. 6a. edición. Santiago, Chile Universitaria
Dourojeanni M (2022) ¿Es posible detener la deforestación en la Amazonia Peruana. In: Castro A, Merino-Gómez YMI (eds) Desafíos y perspectivas de la situación ambiental en el Perú. En el marco de la conmemoración de los 200 años de vida republicana. INTE-PUCP, Lima
Fernandez DP, Neff JC, Huang C, Asner GP, Barger NN (2013) Twentieth century carbon stock changes related to Piñon–Juniper expansion into a black sagebrush community. Carbon Balance Manag 8:13. https://doi.org/10.1186/1750-0680-8-8
Guardiola MDE, Vale Júnior J, Silva EE, Rossi CQ, Pereira MG (2017) Chemical and physical fractions of soil organic matter under various management regimes in Roraima. Braz Semin Cienc Agrar 38:2419–2432. https://doi.org/10.5433/1679-0359.2017v38n4Supl1p2419
Hao X, Ball BC, Culley JLB, Carter MR, Parkin GW (2007). Soil density and Porosity. Soil sampling and Methods of Analysis. CRC Press 2:745–752
Holdrige LR (1967) Life zone ecology. Tropical Science Center, San Jose
Hooke RL, Mart ın-Duque JF (2012) Land transformation by humans: a review. GSA Today 22:4–10
Ibrahim M, Chacón M, Cuartas C, Naranjo J, Ponce G, Vega P, Casasola F, Rojas J (2007) Almacenamiento de carbono en el suelo y la biomasa arbórea en sistemas de usos de la tierra en paisajes ganaderos de Colombia, Costa Rica y Nicaragua. Agroforestería En Las Américas 45:27–36
INEI-Instituto Nacional de Estadística e Informática (2012). IV censo nacional Agropecuario 2012. Perfil Agropecuario Cajamarca. Recuperado de. http://zeeot.regioncajamarca.gob.pe/sites/default/files/mapoteca_virtual/CENAGRO_2012/Guias/CAJAMARCA,2
Junet A, Basile-Doelsc I, Borschneck D, Masion A, Legros S, Marol C, Derenne S (2013) Characterization of organic matter from organo-mineral complexes in an Andosol from Reunion Island. J Anal Appl Pyrol 99:92–100. https://doi.org/10.1016/j.jaap.2012.10.020
Lima AMN, Silva IRD, Neves JCL, Novais RFD, Barros NFD, Mendonça EDS, Demolinari MDSM, Leite FP (2008) Frações da matéria orgânica do solo após três décadas de cultivo de eucalipto no Vale do Rio Doce-MG. Rev Bras Ciênc Solo 32:1053–1063. https://doi.org/10.1590/S0100-06832008000300014
Loss A, Ribeiro EC, Pereira MG, Costa EM (2014) Atributos físicos e químicos do solo em sistemas de consórcio e sucessão de lavoura, pastagem e silvipastoril em Santa Teresa. ES Biosci J 30:1347–1357
Martinkoski L, Vogel GF, Jadoski SO, Watzlawick LF (2017) Qualidade física do solo sob manejo silvipastoril e floresta secundária. Floresta e Ambiente 24:1–9. https://doi.org/10.1590/2179-8087.028216
McLauchlan KK, Hobbie SE (2004) Comparison of labile soil organic matter fractionation techniques. Soil Sci Soc Am J 68:1616–1625. https://doi.org/10.2136/sssaj2004.1616
Morales Ruiz DE, Aryal DR, Pinto Ruiz R, Guevara Hernandez F, Casanova Lugo F, Villanueva Lopez G (2021) Carbon contents and fine root production in tropical silvopastoral systems. Land Degrad Dev 32(2):738–756
Nachtergaele F, van Velthuizen H, Verelst L, Wiberg D, Henry M, Chiozza F, Tramberend S (2023) Harmonized world soil database version 20. Food and Agriculture Organization of the United Nations, Quebec, pp 1–17
Olaya-Montes A, Llanos-Cabrera MP, Cherubin MR, Herrera-Valencia W, Ortiz-Morea FA, Silva-Olaya AM (2020) Restoring soil carbon and chemical properties through silvopastoral adoption in the Colombian amazon region. Land Degrad Dev 32(3720–3730):2020. https://doi.org/10.1002/ldr.3832
Polanía-Hincapié KL, Olaya-Montes A, Cherubin MR, Herrera-Valencia W, Ortiz-Morea FA, Silva-Olaya AM (2021) Soil physical quality responses to silvopastoral implementation in Colombian Amazon. Geoderma 386:114900
Porta J, López M, Toquero C (2003) Edafología para la Agricultura y el medio ambiente, 3rd edn. Mundi-prensa, Madrid
Ríos MM, Dager MR, Carrero J, Tovar MR (2016) Fraccionamiento químico de la materia orgánica en suelos de bosque y sabana. Multiciencias 16:14–21
Rojas J, Ibrahim M, Andrade H (2009) Secuestro de carbono y uso de agua en sistemas silvopastoriles con especies maderables nativas en el trópico seco de Costa Rica. Corp Cienc y Tecnol Agropecu 10:214–223
Roque-Alcarraz R, Silva-Del Águila J, Barrera-Lozano M (2022) Sistemas silvopastoriles para la producción ganadera en el centro de producción limón rocío del UNSM-T. Rev De Vet y Zootec Amazón 2:1–9. https://doi.org/10.51252/revza.v2i2.398
Rucks L, Gracia F, Kaplán A, Ponce J, Hill M (2004). Propiedades Físicas del Suelo. Montevideo, Uruguay. Recuperado de. http://bibliofagro.pbworks.com/f/propiedades+fisicas+del+suelo.pdf
Sanchez CY (2021). Propiedades físicas y químicas de los suelos en sistemas silvopastoriles en Amazonas y San Martín, pp 1–97. https://repositorio.lamolina.edu.pe/handle/20.500.12996/4749
SENAMHI-Servicio Nacional de Metereología e Hidrología del Perú (2023). https://www.senamhi.gob.pe/?p=pronostico-meteorologico
Smith P, House JI, Bustamante M, Sobocká J, Harper R, Pan G, West PC, Clark JM, Adhya T, Rumpel C, Paustian K, Kuikman P, Cotrufo MF, Elliott JA, McDowell R, Griffiths RI, Asakawa S, Bondeau A, Jain AK, Meersmans J, Pugh TAM (2016) Global change pressures on soils from land use and management. Glob Chang Biol 22(3):1008–1028. https://doi.org/10.1111/gcb.13068
Vázquez E, Teutscherova N, Lojka B, Arango J, Pulleman M (2020) Pasture diversification affects soil macrofauna and soil biophysical properties in tropical silvopastoral systems. Agr Ecosyst Environ 302:107083
Vesterdal L, Clarke N, Sigurdsson BD, Gundersen P (2013) Do tree species influence soil carbon stocks in temperate and boreal forests? For Ecol Manage 309:4–18. https://doi.org/10.1016/j.foreco.2013.01.017
Walkley A, Black IA (1934) An examination of the Degtjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci 37:29–38
Winkler K, Fuchs R, Rounsevell M, Herold M (2021) Global land use changes are four times greater than previously estimated. Nat Commun 12:2501. https://doi.org/10.1038/s41467-021-22702-2
Acknowledgements
The authors would like to express their appreciation to the USAID for funding the PEER project “Improving sustainability and resilience of Peruvian Amazon systems through silvopastoralism” and to the producers who provided information and allowed collect data from their farms to completion of this study.
Author information
Authors and Affiliations
Contributions
R.S.M.Conceptualization; Methodology; Data analysis; Writing, review & editing; J.A.O.Conceptualization, Methodology, Review & Editing; D.P.P.Review & editing, Methodology; A.D.Review & editing, Methodology; C.V.G.Data analysis; Writing, Review & Editing; C.G.B.Conceptualization; Review & Editing;
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Salazar, R., Alegre, J., Pizarro, D. et al. Soil carbon stock potential in pastoral and silvopastoral systems in the Peruvian Amazon. Agroforest Syst (2024). https://doi.org/10.1007/s10457-024-00969-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10457-024-00969-w