


Abstract
Permafrost regions, characterised by extensive belowground excess ice, are highly vulnerable to rapid thaw, particularly in areas such as the Yedoma domain. This region is known to freeze-lock a globally significant stock of soil nitrogen (N). However, the fate of this N upon permafrost thaw remains largely unknown. In this study, we assess the impact of climate warming on the size and dynamics of the soil N pool in (sub-)Arctic ecosystems, drawing upon recently published data and literature. Our findings suggest that climate warming and increased thaw depths will result in an expansion of the reactive soil N pool due to the larger volume of (seasonally) thawed soil. Dissolved organic N emerges as the predominant N form for rapid cycling within (sub-)Arctic ecosystems. The fate of newly thawed N from permafrost is primarily influenced by plant uptake, microbial immobilisation, changes in decomposition rates due to improved N availability, as well as lateral flow. The Yedoma domain contains substantial N pools, and the partial but increasing thaw of this previously frozen N has the potential to amplify climate feedbacks through additional nitrous oxide (N2O) emissions. Our ballpark estimate indicates that the Yedoma domain may contribute approximately 6% of the global annual rate of N2O emissions from soils under natural vegetation. However, the released soil N could also mitigate climate feedbacks by promoting enhanced vegetation carbon uptake. The likelihood and rate of N2O production are highest in permafrost thaw sites with intermediate moisture content and disturbed vegetation, but accurately predicting future landscape and hydrology changes in the Yedoma domain remains challenging. Nevertheless, it is evident that the permafrost-climate feedback will be significantly influenced by the quantity and mobilisation state of this unconsidered N pool.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 4.0 license. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
Regions with a high amount of belowground excess ice are especially prone to rapid thaw and associated deep mobilisation of sedimentary organic matter (OM). A prime candidate for rapid and deep thaw processes is the focus region of this study: the Yedoma permafrost domain in Siberia and North America. This permafrost region, covering 2587 000 km2 (Strauss et al 2021) consist of tens of metres of ice-rich silty soil intersected by ice wedges that developed in tundra-steppe environments of the late Pleistocene (as seen from the cliff shown in figures 1 and 2). Other deposits, such as thermokarst lake and drained lake sediments or Holocene cover layers started developing due to permafrost degradation and aggradation during the Late Glacial and the Holocene warm periods (Schirrmeister et al 2020). Because of the region's relatively high OM content and substantial sedimentary volume, the Yedoma domain contains 327–466 Gt organic carbon (OC) and 37–45 Gt total nitrogen (N) (Strauss et al 2017, 2022), which amounts for 26% and 42% of the C and N stocks located in permafrost worldwide, respectively. The thawing of the Yedoma domain can mobilise the carbon reservoirs and release significant amounts of the greenhouse gases carbon dioxide (CO2) and methane (CH4) (Jongejans et al 2021, Schuur et al 2022). It can also enhance nutrient mobilisation, including a primary nutrient N. This is important, since despite the high abundance of N in soil OM, its availability in reactive forms is limited by low temperatures and slow mineralization rate in Arctic soils (Nadelhoffer et al 1991). The increased mobilisation of N due to thawing can promote primary productivity in both terrestrial and aquatic ecosystems (Sanders et al 2022). On the other hand, it can enhance soil decomposition processes, potentially counteracting the benefits of photosynthetic C-uptake by enhanced vegetation growth (Mack et al 2004). Consequently, alterations in N availability can influence the OC balance in the Yedoma domain through these feedback mechanisms, although the overall impact is uncertain.

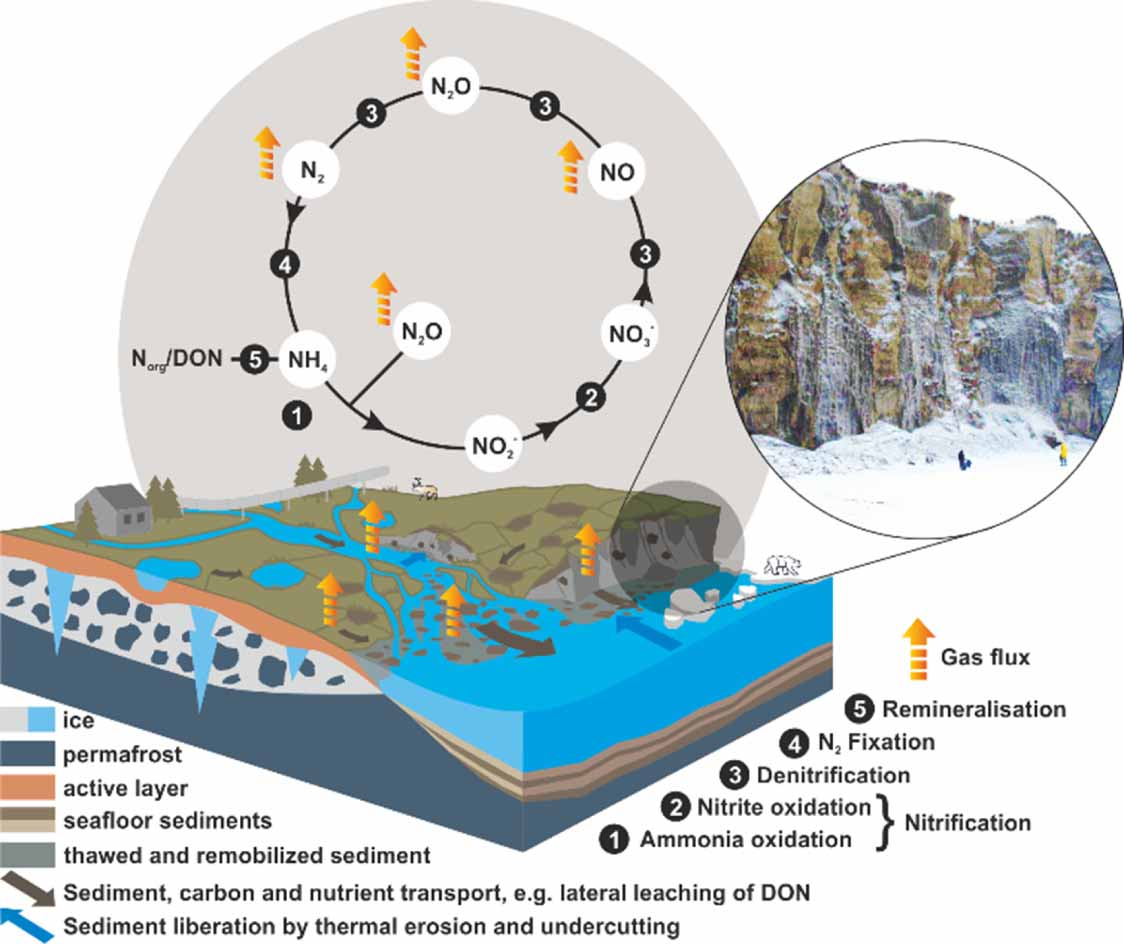
Figure 1. The simplified nitrogen (N) cycle in the context of the potential response of the Yedoma domain (land, river, and delta) to climate change. Terrestrial permafrost thaw in the ice-rich Yedoma domain causes landscape collapse and can affect terrestrial gas fluxes, or it can be leached into freshwaters, affecting its reactivity. The cliff picture inset shows an eroding Yedoma cliff in Alaska at the Itkillik River, Alaska (located ∼69.567, −150.869). Sketch adapted from Mann et al (2022). CC BY 4.0.
Download figure:
Standard image High-resolution image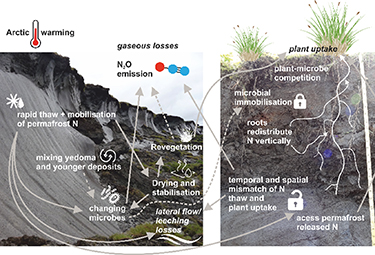
Figure 2. Illustration of alternative usage and loss pathways (gaseous losses, lateral flow/leaching, plant uptake) in various conditions. Background pictures from the Buor Khaya peninsula, Siberia (located ∼71.60, 132.23).
Download figure:
Standard image High-resolution imageIn this study, motivated by the discovery of a significant N stock in the Yedoma domain (Strauss et al 2022), we discuss three main questions related to the fate of soil N, increasingly liberated from the frozen ground in the thawing Yedoma domain: (1) how does the thawed N become accessible for plants and microbes? (2) What are the most likely N-loss pathways through which the liberated N becomes inaccessible to these organisms?, and (3) which are the potential climate feedback loops associated with these different pathways of N uptake and release?
To address these questions we also develop a concept of N-mobilisation, where we include thaw, physical erosion processes, leaching and microbial transformations.
2. N quantity and quality: soil N forms in permafrost and N bioavailability
The ecosystem N cycle consists of multiple microbial and plant-related processes, which are responsible for transformations between numerous solid, soluble and gaseous forms of N (figure 1). The reactive N forms available for biological organisms include mineral N in the form of ammonium (NH4 +), nitrite (NO2 −) and nitrate (NO3 −), as well as dissolved organic N (DON). From these reactive N forms, DON was found to be the most important in high-latitude soils because of the low abundance of mineral N (figure 1) (Jones and Kielland 2002, Fiencke et al 2022). DON comprises approximately half of total dissolved N stocks in a boreal wetland (Kane et al 2010), which is a dominating ecotype of the Yedoma domain, especially in its southern parts.
In the ice-rich Yedoma domain, permafrost warming will increase the size of the active soil N pool due to an increase in seasonally thawed soil volume (figure 2) and more rapid N-cycling processes in warming soil (Schuur et al 2022). From a geomorphological perspective, the two processes determining the fate of N currently locked to permafrost are active layer deepening (gradual thaw), and rapid thaw processes, like thermokarst or thermo/thaw slump erosion (figure 3). This physical process of permafrost degradation is the first step of N-mobilisation through liberation from thawed soil, which needs to happen before any organisms can access the permafrost N. Although thermokarst and erosion caused by excess ice thawing are more prone to happen in ice-rich permafrost such as the Yedoma domain, gradual thaw is happening across the entire permafrost region, encompassing millions of square kilometres beyond the Yedoma domain, where active layers are deepening. This makes N liberation via gradual thaw a phenomenon spanning the circum-Arctic region

{kind=link}
{kind=link}
Figure 3. Simplified visualisation of the Yedoma domain degradation processes with potential of N2O release. Key players are the availability of water, vegetation and microbes (Voigt et al 2020, Fiencke et al 2022). Nitrate (NO3 −), ammonia (NH4 +) as well as dissolved organic nitrogen (DON) are used qualitatively (not quantitatively!) for illustrating mobilisation. As an example: for freshly thawed Yedoma there is a high water-filled pore space/absence of microbial community, and thus a negligible N2O flux potential, although there is freshly thawed N around.
Download figure:
Standard image High-resolution image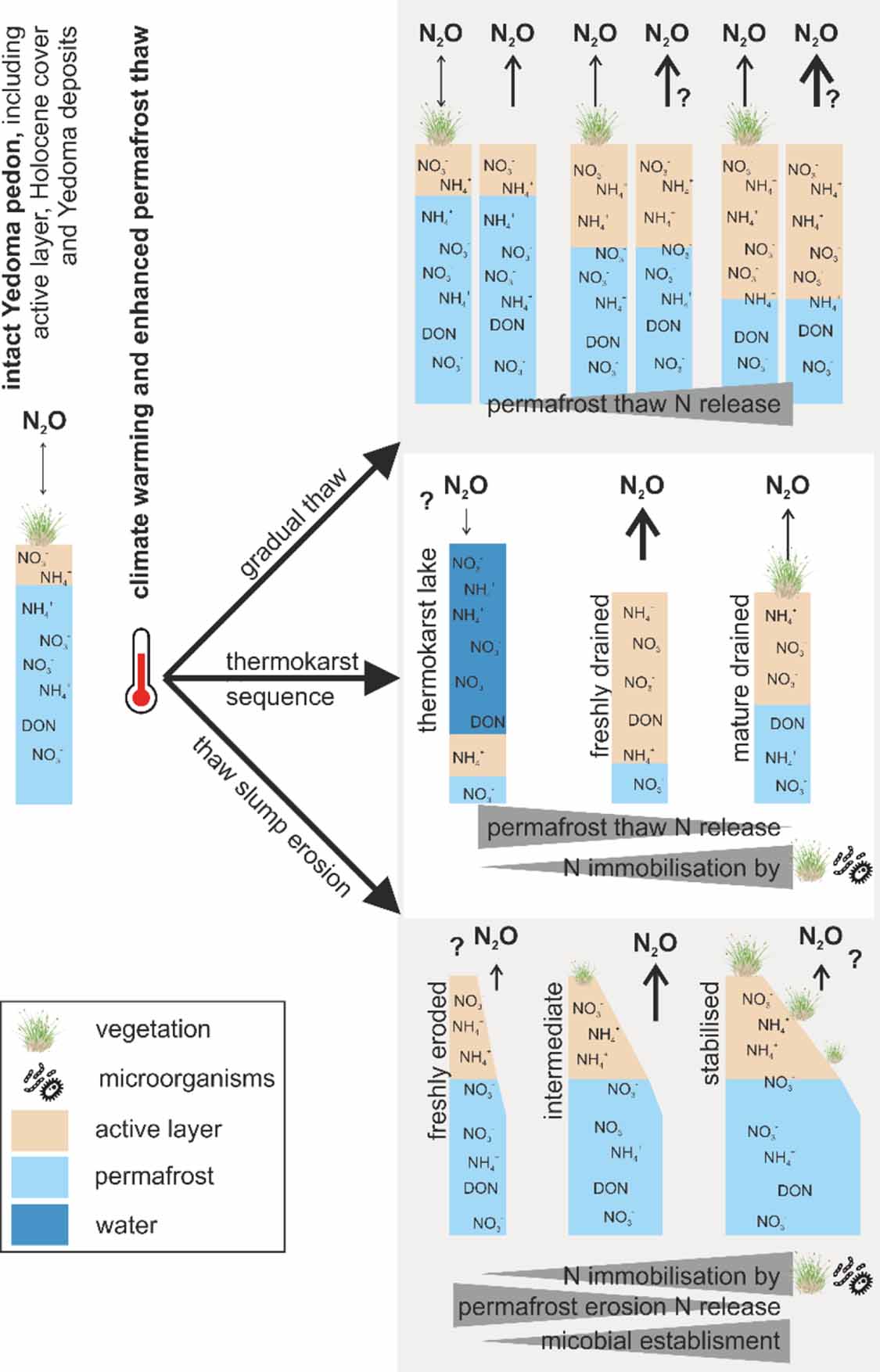{kind=link}
Studies on lability of N compounds in Yedoma deposits are sparse, but one of these rare studies, conducted in Siberian Yedoma deposits (Mack et al 2010), found a liberation of 250 ± 107 µg N g−1 soil yr−1 in a laboratory incubation experiment. Based on these N release rates, they assumed that thaw and decomposition of a 1 cm deep layer of frozen Yedoma deposits alone, if exposed to unfrozen, oxygenated conditions, could supply 3 g N m−2 (Mack et al 2010) to microbial growth and metabolism as well as to vegetation growth. This amount is comparable to plant N uptake rates of 0.2–8.3 g N m−2 yr−1 reported for shrubby grass, tussock moss and forest tundra vegetation, summarised by Wild et al (2018), clearly demonstrating the importance of this added N liberated from permafrost. Additionally, significantly higher rates of net N mineralisation and nitrification were found in thawed Yedoma as compared to Holocene cover deposits (Marushchak et al 2021).
Differences in N density between the Yedoma domain's active layer (1.6 kg N m−3) and the Yedoma domain's permafrost soils (0.9–2.2 kg N m−3) were found to be small (Strauss et al 2022). While Arctic soils contain significant N stocks in soil OM, typically little of this N is biologically available (Shaver et al 1992, Buckeridge et al 2010, Beermann et al 2017). The fraction of readily biologically available dissolved N in Arctic ecosystems ranges from 0.01% to 3% of total soil N stocks (Shaver et al 1992, Buckeridge et al 2010).
A simple, first order proxy for the bioavailability of N following thaw is the C:N ratio. Compared to Pleistocene (Yedoma) and early Holocene permafrost deposits (Taberites), the active layer soils of the Yedoma domain were found to have higher C:N ratios (Strauss et al 2022), implying a lower rate of N mineralisation there (Booth et al 2005, Liu et al 2017). Significantly higher rates of net N mineralisation and nitrification were found in thawed Yedoma as compared to Holocene cover deposits (Marushchak et al 2021). An accumulation of NH4 + was observed within Yedoma permafrost as compared to the active layer (Strauss et al 2022), likely due to mineralisation of OM with lower C:N ratios and delayed or lacking plant uptake (Uhlířová et al 2007). This elevated supply of NH4 + could also fuel the nitrification rates. Recently, Hansen and Elberling (2023) confirmed this by stating that permafrost soils across the Northern Hemisphere contain larger concentrations of biologically available inorganic N (mainly as NH4 +) than the active layer. Also in sub-Arctic peatlands, available N in near surface permafrost were seven times higher than in active layer permafrost; and higher rates of net N mineralisation and plant N uptake were also observed (Keuper et al 2012). For permafrost from the Tibetan Plateau, the opposite case was found, having more N in the active layer than in the permafrost below (Mao et al 2020).
The vegetation type dominating during the time Yedoma deposits accumulated can partly explain the high lability of N compounds in Yedoma permafrost. Most of the Yedoma domain includes thick permafrost deposits that accumulated in the late Pleistocene, when grassland ecotype was dominating there (Zimov et al 2012). In their synthesis study on gross N mineralisation rates, Booth et al (2005) found that OM from grasslands appears to have lower C:N ratios and is inherently more productive for NH4 + than OM from forested or woody sites with higher C:N ratios. The mobilisation of the deeper still perennially frozen N rich Yedoma OM during permafrost thaw likely represents an important source of additional bioavailable N to the terrestrial as well as adjacent aquatic ecosystems.
3. Fate of N from thawed permafrost
3.1. Plant N uptake
Since plant growth in high-latitude ecosystems is usually N limited (Beermann et al 2015, 2017), N release from thawing permafrost deposits may boost plant productivity and thus CO2 fixation from the atmosphere through photosynthesis. However, the strength of this feedback is strongly affected by how well the liberated nutrients including N will be accessible for plants (Pedersen et al 2020) or microbes (Baker et al 2023). The accessibility of nutrients is influenced by the spatial and temporal patterns of N release and transport, the presence or absence of plants, plant, species composition, plant rooting depth as well as soil characteristics. Seasonal variability plays an important role here: Plants and microbes are able to immobilise available N (figure 2) before photosynthetic activity sets in during the winter/spring transition (Edwards and Jefferies 2010). However, seasonal declines in microbial biomass may result in another pulse of available N (Bardgett et al 2007). Active layer deepening, apart from exposing more OM to microbial decomposition, leads to a longer thawing period, thereby prolonging the period with biologically available soil N.
Active layer deepening likely also results in direct N inputs to the ecosystem N cycle from newly thawed permafrost to the bottom of the active layer, as well as longer soil N availability, especially in the first years after thaw (Salmon et al 2016, 2018).
It has been shown, however, that due to a temporal mismatch between peak vegetation growth (mid-summer) and maximum thawing depth (late summer/autumn), there is incomplete utilisation of nutrients by plants near the permafrost table (Lacroix et al 2022). Also, plants will not necessarily be able to access the N released from permafrost, if they are located deeper down than their rooting zone (Keuper et al 2017). Incomplete plant N use can cause a nutrient oversupply near the permafrost table, which may lead to freely available N for microbial cycling processes such as nitrification and denitrification. This, in turn, could lead to emissions of N2O, a strong greenhouse gas and stratospheric ozone-depleting substance (figure 2 and Lacroix et al (2022)), or increased losses of N via downward vertical leaching and lateral flow (figure 2, chapter 3.2). This may be the case with gradual active layer deepening, while abrupt thaw exposes the newly thawed material to the surface, making it readily available for newly established vegetation on retrogressive thaw slumps (Marushchak et al 2021) or in thermokarst lakes (Walter Anthony et al 2014).
In retrogressive thaw slumps and other disturbed sites, however, development of full vegetation cover takes several years, and easily exceeds the capacity of plants to take up N during early stages of succession the mineral N supply (Marushchak et al 2021). This successional mismatch is probably also true for the process of surface water generation resulting from the melting of ground ice and subsequent subsidence, although there is limited literature on this topic. Thermokarst lake and gully formation leads to distinct vegetation dynamics, influencing the utilisation of newly available N by plants. The development of aquatic landscape features, such as small pools and troughs, requires colonisation by aquatic vegetation (primarily consisting of mosses and sedges) before they can effectively utilise the accessible N for carbon sequestration. Nevertheless, once the full vegetation cover is established on these disturbance sites, they exhibit the potential for considerable productivity.
Thus, increased N availability by permafrost thaw in soils could enhance net primary productivity and increase OC stocks in plant biomass, but this might not always be the case. Some experimental evidence confirms that plant growth increases with newly thawed deep N sources, while other studies show that effects on OC acquisition might be limited since only few deep-rooting plant species will be able to exploit the newly thawed N (Keuper et al 2017). Considering that a large part of the Yedoma domain lies within the continuous permafrost zone (with a shallow active layer depth of <1 m), it is likely that N in the soil layers exposed by active layer deepening may become accessible to deep rooting plants (see the gradual thaw pathway in figure 3), if there is no temporal mismatch.
Recently, a study of vegetation types in retrogressive thaw slumps (Buckeridge et al 2023) revealed that shrubs are the best candidate for plant N uptake as they have a deep vertical rooting potential (figure 2) and can reach the deep N stocks, as also found by Hewitt et al (2019). This is in line with the shrubification which has been observed in Arctic tundra due to recent warming (Frost et al 2013). Whether increased vegetation stocks may offset soil OC losses is therefore still uncertain (McGuire et al 2018, Mauritz et al 2021, Schuur et al 2021, 2022).
3.2. Lateral and vertical flow
In the Yedoma domain (and other ice-wedge-bearing permafrost), the thawing of ground ice frequently results in elevated surface water levels. This phenomenon becomes apparent through the degradation of ice-wedge polygonal landscapes, resulting in the formation of polygonal pools and ice wedge troughs, and sometimes leading to interconnected hydrological pathways (Liljedahl et al 2016). Importantly, this process is likely to serve as an increasingly significant source of nutrients (Vonk et al 2015). For instance, Koch et al (2018) observed a rapid decline in ammonium (NH4 +) levels within deep trough ponds, attributable to biogeochemical cycling.
Runoff from Holocene cover on top of Yedoma uplands and some refrozen thermokarst basins sediments of the Yedoma domain can transport nutrients like NO3 − and NH4 + to aquatic systems. This runoff/lateral flow is limited in deeper strata, especially in Taberites beneath thermokarst lakes, as they are (at least partially) (re-)frozen. Vertical flow could occur as well if a groundwater connection is present, or through transport within plant roots in a through-going talik. Particulate N and DON originating from runoff could be deposited on floodplains, in river deltas, or in the nearshore sediments of the Arctic shelf seas (Ramage et al 2018, Marzadri et al 2021). If leached and transported to the aquatic systems, the fate of N strongly depends on the form and quality of N, as well as other stream network characteristics such as redox potential, oxygen status, availability of other nutrients, pH, temperature and microbial and plant community composition. Additionally, water transparency and sediment content are controlling the availability of light and thus nutrient uptake and primary productivity.
Nutrient losses through lateral flow and vertical leaching are potentially a major factor in overall tundra nutrient budgets (Mack et al 2004, Schaeffer et al 2013). Given the observed increases in tundra stream NO3 − export from 1978 to 2006 (McClelland et al 2007, Townsend-Small et al 2011), understanding mechanisms for N availability and losses in tundra soils is important. However, estimates of nutrient leaching losses from tundra soils remain limited (Harms and Jones Jr 2012, Treat et al 2016). Rates of N mineralisation, N assimilation and immobilisation vary seasonally due to interactions with plant biomass and microbes (Schmidt et al 2002, Sorensen et al 2008, Baker et al 2023) and further complicate estimates of N leaching losses. For example, soil N losses increased significantly during the late growing season at a high-Arctic tundra site as plants senesced (Schaeffer et al 2013).
Losses of DON account for the majority of soil N leaching to streams from pristine ecosystems (Hedin et al 1995, Perakis and Hedin 2007) like Arctic ecosystems (Lafrenière and Lamoureux 2008, McNamara et al 2008). The export of dissolved OM, and therefore DON, is related to dissolved OM concentrations within the soils, dissolved OM retention within the soil, and hydrologic flux from the soil.
The export of dissolved OM is also controlled by the amount of runoff. Olefeldt and Roulet (2012) found substantially lower dissolved OM fluxes from a permafrost peatland than from other peatlands and concluded this was the result of less runoff due to presence of impermeable permafrost. Changes in the flow path of water due to permafrost thaw will alter dissolved OM and N inputs to aquatic ecosystems, resulting in predictions of increasing DON fluxes in Siberian watersheds (Frey et al 2007).
In streams, nutrient export is highly variable over time and space. The majority of nutrient export in Arctic rivers occurs during spring freshet (Holmes et al 2012, Juhls et al 2020). Holmes et al (2012) estimated dissolved N loading to be 1.26 Tg per year; roughly two-thirds (0.85 Tg) was organic N over the inorganic remainder (NO3 − and NH4 +). Additionally, there is seasonal variability in the quantity of the dominant N form released in the Yedoma domain. As was found by Sanders et al (2022), who studied the Yedoma domain, DON is predominantly transported to the ocean in summer, while the concentrations of exported DON and NO3 − are similar during periods with ice cover.
Besides quantity, the quality of dissolved OM is also temporally variable. OM quality differs during the spring freshet from the growing season; 20%–46% of dissolved OM lost during spring freshet is degraded relatively rapidly (Michaelson et al 1998, Holmes et al 2008), while dissolved OM exported to the Arctic ocean during the summer is slow to degrade (Holmes et al 2008). N released by dissolved and particular OM mineralisation may result in increased stream productivity or N retention (Peterson et al 2001), denitrification (Peterson et al 2001), or export to the Arctic Ocean (Frey et al 2007, Mann et al 2022, Sanders et al 2022, Ogneva et al 2023). The net effect on climate, considering direct and indirect greenhouse gas emissions from both OC and N cycles is presently difficult to estimate from lakes and rivers in the Yedoma domain, and a solid estimation for N2O emissions from Arctic rivers is lacking until now.
3.3. Microbial immobilisation and changes in decomposition because of improved N availability
Excess N liberated during thaw will be partly immobilised by being incorporated into the microbial biomass following thaw, but with continued supply of N, the likelihood of excess N availability increases. This excess N is, depending on the micro-conditions, simultaneously available for various microbial transformation processes including N2O and molecular N (N2) production as long as growth of vascular plants remains hampered (Palmer et al 2012, Palmer and Horn 2012, Hetz and Horn 2021, Horn and Hetz 2021).
The effect of excess N is a subject of discussion, giving rise to two distinct phenomena: the concept known as the 'N mining theory' (Lovett et al 2018) and the 'N priming effect' (Mack et al 2004). The N mining theory suggests an increase in decomposition due to N limitation, where microbes are forced to mine for additional N-substrates from OM. The theory suggests that when N is limited, N-acquiring microbes can use OC as an energy source, thereby enhancing OM decomposition and increasing respiratory CO2 losses to acquire the needed N (Chen et al 2014, Lovett et al 2018). The N priming effect, on the other hand, can be positive or negative, entailing a boost or a decrease in overall OM decomposition through enhanced N availability.
Notably, the N-priming effect not only enhances the decomposition of OM and stimulates further N-release, but also leads to higher N availability and enhanced decomposition of OM increase OC release (Mack et al 2004, Lavoie et al 2011). For example, Mack et al (2004) showed that aboveground OC storage increased following N-addition, but was offset by increased OC decomposition—and thus soil OC losses—belowground. Similarly, Zhang et al (2023) found that a warming-induced, enhanced soil N supply stimulated soil microbial activity and amplified soil OC losses from permafrost soils in the Tibetan Plateau. These findings support the 'stoichiometric decomposition theory', which suggests that N addition is beneficial for OM decomposition.
In contrast to these findings, Berg and Matzner (1997) and Riggs et al (2015) showed that increased N availability could lead to decreased OM decomposition and thus negative priming. Riggs and Hobbie (2016) hypothesised that negative priming can be caused by three mechanisms. First, increased N availability can improve OC use efficiency leading to OC use for microbial growth rather than respiration (Weintraub and Schimel 2003, Manzoni et al 2012, Chen et al 2018). Second, N addition might lead to decreased activity of oxidative enzymes, thereby limiting decomposition of complex OC molecules such as lignin (Eisenlord et al 2013). Third, soil acidification following N addition might lead to decreased microbial biomass and hence to decreased OM decomposition rates (Mueller et al 2012).
For Alaskan Arctic and boreal soils, Lavoie et al (2011) found that N addition could have a positive as well as a negative effect on OM decomposition. They showed that a positive effect was mainly observed for organic soils, whereas negative effects were mainly found for mineral soils, from which they concluded that the direction of priming in decomposition is highly dependent on the soils' labile OM fraction. Likewise, Wologo et al (2021) found that addition of N (in the form of NH4 + and NO3 −) and acetate, compounds abundant in permafrost including Yedoma (Mann et al 2022), triggered the breakdown of some dissolved OM compounds, but resulted in a net decrease in overall decomposition rate. Thus, depending on the OM degradability and the microbial composition, the N stock in the Yedoma domain might pose either a positive or a negative climate feedback upon thaw.
4. Newly thawed permafrost N: a positive or negative climate feedback?
4.1. Likelihood for a negative feedback
Permafrost thaw will increase the amount of bioavailable N in an N-limited environment, which could therefore boost ecosystem productivity resulting in a negative climate feedback. Walter Anthony et al (2014) suggest that N and other nutrients such as P released from thawing Yedoma deposits have supported high lacustrine primary production and OC accumulation in thermokarst lake sediments during the Late Glacial to Holocene warming. In addition, in re-vegetated (stabilised) thermokarst basins, plants will be able to access former deep Yedoma N (see figure 3).
On the one hand, as mentioned before, abrupt thaw could increase OC sequestration, while gradual thaw poses the potential for incomplete nutrient use by plants (due to temporal and spatial mismatch). On the other hand, rapid thaw processes disturb or destroy the plant cover and thereby hinder the possibility of plants to catch the initial thaw liberates nutrient pulse. Therefore, it is possible that a considerable amount of N is not used to promote OC sequestration in the short-term. As a result, the possibility to create a negative feedback to climate via enhanced plant growth is limited in these regions with on-going thermokarst activity.
4.2. Likelihood for a positive climate feedback by gaseous losses
While part of the excess N liberated from thawing permafrost in the Yedoma domain will be used for plant growth and transformed into microbial biomass with continued N supply as described above (figure 2), part of it may become available for microbial transformation processes such as nitrification and denitrification (Horn and Hetz 2021, Ramm et al 2022). These microbial N transformation processes may cause gaseous N losses in the form of N2O as well as N2. N2O is produced mainly by denitrification but also as a side product during the first step of nitrification, the ammonia oxidation. Further, nitrification has a crucial role for denitrification via NO3 − supply to serve as the electron acceptor in denitrification. Recently published data on high N2O emissions and potential N2 losses from retrogressive thaw-slumps within Yedoma deposits demonstrate that this is a likely scenario for such permafrost thaw features in the Yedoma domain (Marushchak et al 2021).
There is increasing evidence from soils across the permafrost region that, despite their general N limitation due to slow N mineralisation in cold and often humid conditions, permafrost-affected soils can be substantial sources of N2O when three conditions are met (Repo et al 2009, Voigt et al 2017b, 2020, Marushchak et al 2021). First, a sufficient mineral N supply is a key factor controlling N2O emissions from soils (Parton et al 1996). In this context, the C:N ratio has been linked to N2O emissions from soils with highest emissions occurring at low to intermediate range C:N ratios, which support N mineralisation (Klemedtsson et al 2005, Liimatainen et al 2018). In the Yedoma domain, the C:N ratios ranged between 9–19 (medians of the different compartments in Strauss et al (2022), which is low enough to support N2O emissions as shown in Marushchak et al (2021).
Second, a lack of vegetation growth increases the mineral N availability for microbes and is thus favourable for N2O emissions. Previous studies reported high N2O emissions in locations where vascular plants are absent or N uptake by vegetation is otherwise disturbed, e.g. by thermokarst processes (Marushchak et al 2011, Gil et al 2017, Voigt et al 2017a). This is frequently the case in the Yedoma domain with its high abundance of eroded surfaces, disrupted plant growth, and disturbed site conditions due to abrupt thaw processes.
Third, an intermediate moisture content has been shown to promote N2O emissions from permafrost-affected soils (Voigt et al 2020). On the one hand, water-saturated conditions hinder N mineralisation and nitrification, which are aerobic processes, thereby limiting provision of mineral N species used for N2O production. At the same time, a high water content promotes denitrification as this is an anaerobic process. If denitrification is complete under water-saturated conditions, N2O is reduced to the inert N2 gas, and complete denitrification may also lead to atmospheric N2O uptake (Butterbach-Bahl et al 2013, Voigt et al 2017b, 2020). On the other hand, an intermediate moisture range allows both NO3 − and N2O production via aerobic nitrification as well as N2O production via anaerobic denitrification to co-occur (Butterbach-Bahl et al 2013, Abbott and Jones 2015, Fiencke et al 2022). Thus, the magnitude of N2O emissions following permafrost thaw will largely depend on hydrology.
Limitations in microbially available OM together with accumulation of NO3 − associated with high nitrification rates might further enhance N2O production during denitrification (Hetz and Horn 2021). In favourable, intermediate soil moisture conditions, a microbial community capable of intensive mineral N cycling (by nitrification and denitrification) and substantial N2O production can be established in these mineral N rich sediments (Marushchak et al 2021). While for freshly thawed, wet Yedoma sediments, low N2O emissions were found due to low abundance of key functional groups, particularly nitrifiers, whereas high N2O emissions occurred after a few years of stabilisation, drying, partial re-vegetation and changes in microbial community (Marushchak et al 2021).
Based on these initial findings, we thus expect that the following areas are prime candidates for substantial N2O release (figure 3): thaw slumps, freshly drained lake basins, and thermo-erosional valleys. If thermokarst lakes drain, unfrozen sub-lake taliks can turn into zones of intermediate soil moisture levels. Nitze et al (2020) observed widespread lake drainage in the Yedoma domain over the past decades and predicted a similar trend of more lake drainage events in the future, especially for southern margins of continuous permafrost. With improved drainage, the OM in lake sediments is exposed to enhanced mineralisation, which will boost mineral N production and the microbial N transformation processes releasing N2O.
5. Estimating the future net climate feedback from newly thawed N
The net effect on the climate of newly thawed N from permafrost degradation in the Yedoma domain strongly depends on local conditions (e.g. prevailing wetness, vegetation, microbes). These conditions will determine whether N in thawing permafrost is taken up by vegetation or microbes or if it will be subjected to vertical leaching or to microbial transformation processes, such as denitrification, leading to N losses in the gaseous form. In the short term, the gradual deepening of the active layer will affect only a small portion of the currently frozen N pool in the Yedoma domain. Most of this newly thawed N of the newly thawed active layer will be at depths interacting with plants, thus with a potential for enhancing the carbon sink capacity of these regions and limiting losses though lateral runoff and gaseous, vertical losses such as N2O emission. Speculative, but possibly further long-term climate warming beyond the 21st century will lead to an increasingly thawed N pool at depths that are below the root reach of plants. Even at present, a temporal mismatch of N supply and demand can leave part of the mobilised N inaccessible for plants. Furthermore, thermokarst lake initiation, coastal and riverbank erosion as well as erosion on thermokarst basins will continue to thaw deep Yedoma deposits, thereby mobilising substantial amounts of deep N below 3 m. The N at these soil depths could become available for microbes participating in N cycling under intermediate soil moisture levels, which might lead to N2O production.
Another difficulty for estimating N2O production based on environmental conditions is related to the process of stabilisation of rapid thaw (for example in thaw slumps and cliff erosion), which seems to be decoupled from hydrology parameters driving the N2O production elsewhere. Freshly thawed Yedoma from a permafrost exposure in Siberia was found to be inert for N2O production right after thaw with gradual but significant increase in N2O emissions within the first few years after thaw (Marushchak et al 2021). This temporal dimension of the already highly spatially variable N2O production in the landscape further increases the uncertainty for the estimation of the future net climate feedback in upscaling attempts. Similarly, Chen et al (2018) found that at the early stage of permafrost collapse, microbial N limitation is cancelled out because of enhanced microbial N mineralisation. This results in a lower C:N ratio and high N availability, which might lead to higher metabolic efficiency, thereby decreasing CO2 release. Later on, due to N leaching or consumption, N may become again limited and CO2 release will likely increase. This illustrates that these processes probably happen on different time scales. Besides these temporal complexities, uncertain future shifts in soil moisture conditions can crucially alter N2O production rates.
To wrap up the future the Yedoma domain pathway: There will be substantial mobilisation of N by deep permafrost thaw. This, in addition to the N release by gradual deepening of the active layer could provide a substantial amount of additional N. According to a simplistic back-on-the-envelope calculation approach by Ramm et al (2020), we can illustrate the significance of thaw enhanced N cycling and N2O release in the Yedoma domain. If 10% (following the projected 9.6% Yedoma carbon loss over the 50 incubation year period from Strauss et al (2017)) of the N stored in the Yedoma domain (4.1 Gt, Strauss et al (2022)) is released between the present day and the year 2100 (assumption based on Schuur et al (2022); Schuur et al (2015)), and 1% (IPCC's default N2O emission factor from mineral soils) is emitted as N2O (0.4 Tg N2O-N yr−1, 31 Tg N2O-N until 2100), this would be equivalent to ∼6% of the global annual rate of N2O emissions from soils under natural vegetation (6.6 Tg N2O-N yr−1 (Ciais et al 2013) already. For the total permafrost N stock of 97 Gt (Harden et al 2012, Strauss et al 2022), this would amount to 1.0 Tg N2O-N yr−1, 76 Tg N2O-N in total for the 76 years to 2100 and 15% of the global annual rate of N2O emissions from soils under natural vegetation, which have not yet been accounted for. The release of N2O resulting from thawing permafrost, particularly Yedoma, is an additional contribution to the current N2O emissions originating from permafrost soils which have been estimated to range between 0.14 and 1.27 Tg N2O-N (Voigt et al 2020). Thus, the thawing Yedoma domain is likely a neglected N2O source. Our study shows potential pathways of N release when we further 'open'the Yedoma-domain-permafrost freezer by human-caused climate warming. Future research should prioritize studying N flow paths post-thaw of permafrost deposits, along with physico-chemical and microbial stabilization/mobilization of these nutrient reservoirs. Additionally, developing process-based models to simulate N and carbon dynamics can enhance predictions regarding permafrost-climate feedbacks in vulnerable ecosystems.
6. Conclusion
The increasing release of N from the recently found, until now largely freeze-locked soil N pools in the Yedoma domain could further accelerate climate feedbacks from permafrost thaw through additional N2O emissions, or it could reduce climate feedbacks through promoting enhanced vegetation C sequestration. While the latter largely depends on how well the spatial and temporal pattern of plant N uptake matches with those of N release, the Yedoma deposits are a prime candidate for N2O production if the conditions are right (i.e./e.g. medium water content, establishment of microbial community capable of active N cycling, partial vegetation cover). More precisely, stabilising permafrost thaw sites with intermediate soil moisture content and disturbed vegetation cover likely have the highest potential for N2O release. However, making reliable predictions of future landscape and hydrology changes in the Yedoma domain remains a major challenge due to lack of data. In all cases, there is strong evidence that the permafrost-climate feedback will be affected by the amount and state of mobilisation of this previously unquantified N pool. Future research should focus on quantifying N flow post-thaw in permafrost and developing process-based models to improve predictions on permafrost-climate feedbacks.
Acknowledgments
This effort is embedded into the Permafrost Carbon Network (PCN). We acknowledge the support by the European Research Council FluxWIN (#851181) and the German Federal Ministry of Education and Research (BMBF; CACOON (#03F0806A), EISPAC (#03F0809A). The work of CB, CV, and MEM was supported by the Academy of Finland/Russian Foundation for Basic Research project NOCA (General Research Grant from the Academy of Finland, Decision No. 314630), the Yedoma-N project (General Research Grant from the Academy of Finland, Decision Number 287469) and N-PERM (General Research Grant from the Academy of Finland, Decision Number 341348). C V was further supported by the Academy of Finland project MUFFIN (Decision No. 332196), and the BMBF project MOMENT (No. 03F0931A), MEM by the Academy of Finland project Thaw-N (Decision No. 349503) and CB by the Lise Meitner Program of the Austrian Science Fund (FWF) with the project PERNO (Grant Number M 3335-B).
Data availability statement
All data that support the findings of this study are included within the article (and any supplementary files).
Contributions
J S designed the study and generated the first draft further improved by M E M, L v D, T S, C B, C V, L L J, and C T. All authors contributed significantly by revising and reviewing the manuscript drafts.
Ethics declarations
The authors declare no competing interests.