
Abstract
Genetically engineered (GE) cotton event MON 88702, producing Mpp51Aa2 (previously mCry51Aa2) from Bacillus thuringiensis (Bt), controls sucking pests, such as Lygus spp. (Hemiptera: Miridae) and thrips (Thysanoptera). Ingesting high doses of the insecticidal protein resulted in adverse effects on life table parameters of beneficial, predatory Orius spp. (Hemiptera: Anthocoridae). This triggered laboratory studies with more realistic food treatments, including different combinations of prey types with and without Bt protein to further characterize risks to this important group of non-target organisms. In this work, exclusive feeding of frozen spider mites (Tetranychus urticae, Acari: Tetranychidae) from Bt cotton confirmed adverse effects on longevity and fecundity of O. majusculus adults. Alternate feeding of Bt protein-containing spider mites and Bt-free Ephestia kuehniella (Lepidoptera: Pyralidae) eggs mitigated effects on longevity, but not on fecundity. When living larvae of Spodoptera littoralis (Lepidoptera: Noctuidae) from Bt cotton were fed to the predators, however, no effects on longevity and reproduction of female O. majusculus were observed, despite the fact that Bt protein concentrations in larvae were almost as high as concentrations in spider mites. When a diverse mix of prey species with various Bt protein concentrations is consumed in the field, it is unlikely that exposure of Orius spp. to Mpp51Aa2 is high enough to exert adverse effects on predator populations. MON 88702 cotton may thus be a valuable tool for integrated management of sucking pests.
Similar content being viewed by others


Introduction
Biotechnology has been deployed to generate crop plants with resistance against major insect pests. Genetically engineered (GE) cotton event MON 88702 produces Mpp51Aa2 (previously mCry51Aa2), a modified insecticidal protein from Bacillus thuringiensis (Bt) (Gowda et al. 2016). While previous Bt cotton products carrying cry1, cry2, or VIP genes target Lepidoptera pests, MON 88702 effectively controls sucking pests, such as Lygus spp. (Hemiptera: Miridae) and thrips (Thysanoptera: Thripidae) (Akbar et al. 2019). In integrated pest management, the avoidance of broad-spectrum insecticides by using more environmentally friendly and selective pest control measures is key so that a rich complex of beneficial species can provide ecosystem services, such as biological control, decomposition, and pollination (Wijnands et al. 2012). Environmental risk assessment, which precedes every release of GE plants, includes an evaluation of potential non-target effects (Romeis et al. 2008). Because MON 88702 mainly targets bugs (Hemiptera, suborder Heteroptera), adverse effects on predatory bugs are of particular concern. In fact, when high doses of Mpp51Aa2 were provided in artificial diet, survival of 5-day-old Orius insidiosus (Say) (Heteroptera: Anthocoridae) nymphs was reduced (Bachman et al. 2017). In addition, spider mites from Bt cotton, which contained relatively high concentrations of Mpp51Aa2, caused impaired survival and development when fed to neonate Orius majusculus (Reuter) (Heteroptera: Anthocoridae) (Kim et al. 2021). Moreover, when 5-day-old nymphs were fed with spider mites from Bt cotton, the emerging females laid approximately 50% fewer eggs than females fed with spider mites from non-Bt cotton (Kim et al. 2021). In more realistic exposure scenarios with alternative prey that contained less or no Bt protein, such as noctuid larvae or cotton aphids, effects on the development of O. majusculus neonates were absent. When spider mites from Bt cotton were fed for limited time periods, effects on nymphs were less pronounced compared with exclusive Bt spider mite feeding (Boss et al. 2023).
The aim of this study was to further characterize potential risks of Mpp51Aa2-producing cotton on beneficial anthocorid bugs by assessing the effects of combinations of different prey types with and without Bt protein. While Boss et al. (2023) focused on juvenile development of O. majusculus, the current study investigated longevity and fecundity of adults. Exclusive feeding with spider mites or noctuid larvae reared on Bt cotton was compared with scenarios where prey from Bt cotton was alternated with flour moth eggs that contained no Bt protein.
Material and methods
Plants and insects
Cotton (event MON 88702 producing Mpp51Aa2, “Bt cotton”) and the non-transgenic near isoline DP393 (“non-Bt cotton”) were obtained from Bayer Crop Science (St. Louis, USA). Plants were cultivated in a climate chamber according to Boss et al. (2023) and used from 6 weeks after sowing. Mpp51Aa2 is the revised, structure-based name of modified Cry51Aa2 (Crickmore et al. 2021).
Tetranychus urticae Koch (Acari: Tetranychidae) spider mites were reared continuously on Bt or non-Bt cotton situated in one large environmental climate chamber (Boss et al. 2023). A preliminary experiment revealed that the number of spider mites needed for the planned experimental setup exceeded the capacity of the culture. Spider mites were thus collected over several months by beating the Bt or non-Bt cotton plants in the climate chamber over a tray and stored at − 70 °C. Instead of live prey, frozen spider mites were used for the here described experiments.
Spodoptera littoralis (Boisduval) (Lepidoptera: Noctuidae) was provided by Syngenta Crop Protection AG (Stein, Switzerland) and reared at Agroscope as described by Meissle et al. (2023). This species, also called cotton leafworm, is an Old World pest of cotton and thus a realistic prey of O. majusculus. Eggs of S. littoralis were collected daily. Every other day in the afternoon, neonates were placed on leaf discs from Bt or non-Bt cotton and incubated at 22 °C. In the next morning, living larvae were used for feeding experiments. The feeding time was kept short to ensure that larvae were still small (first instar) and thus easy to handle for O. majusculus. The green colour of the larvae and microscopic observations confirmed that their gut was filled with cotton tissue.
Alternative Bt protein-free prey consisted of Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) eggs. While this storage pest is not a realistic prey for O. majusculus, it served as a model for Bt-free food that is known to have a high nutritional value for O. majusculus development and reproduction (Bonte et al. 2017; Boss et al. 2023). The species was not reared at Agroscope, but sterilized eggs were ordered from Agroline Bioprotect (Aesch, Switzerland), and stored at − 20 °C. No Bt-treatment was involved in the rearing of the species.
An O. majusculus culture was maintained on E. kuehniella eggs as food and bean pods as egg-laying substrate (Boss et al. 2023). To obtain specimens for experiments, batches of bean pods with fresh eggs (< 24 h) were incubated at 22 or 25 °C. Once eggs had hatched, bean pods were removed and nymphs were fed with E. kuehniella eggs until adult emergence.
Feeding experiments with spider mites
Experiments started with adults that had emerged within the previous 48 h. After O. majusculus were sexed (Ferragut and González-Zamora 1994), one male and one female were introduced to a ventilated plastic dish (5 cm diameter), lined with a moist cotton cosmetic pad and a Bt or non-Bt cotton leaf disc (4 cm diameter). In the first series of experiments, the O. majusculus pairs were assigned randomly to one of five food treatments (Table 1): SM: prey consisted of frozen spider mites from either Bt or non-Bt cotton, presented on the Bt or non-Bt cotton leaf discs, respectively; SM/EE alt.: frozen spider mites from Bt or non-Bt cotton were fed alternately with frozen E. kuehniella eggs in 2-day periods, while half of the replicates started with mites as prey and the other half with eggs; EE: prey consisted of E. kuehniella eggs, presented on either Bt or non-Bt cotton leaf discs; EE/NF alt.: E. kuehniella eggs were fed alternately with no additional food in 2-day periods, while half of the replicates started with eggs and the other half with no additional food; NF: no additional food was provided on the Bt or non-Bt cotton leaf discs. Dishes with O. majusculus were incubated at 25 °C, 70% relative humidity and 16 h light. Every second day, leaf discs, cotton pads, and dishes were changed and new food was provided. In the treatments involving spider mites, new spider mites were added on the days in between, because they dried out quickly. In the treatments with alternating food, the food type was switched with every change of the leaf disc (2-day intervals). Replaced leaf discs with O. majusculus eggs were further incubated and water was added as needed to keep the cotton pads moist. After 4 days, the number of empty eggs (nymph has hatched = viable egg) and the number of closed eggs (either infertile egg or nymph died before hatching) was counted under a binocular microscope. Males dying within the first 5 days of the experiment were replaced to ensure insemination of females. The experiment was terminated after 24 days. Recorded parameters were survival of males and females (daily), preoviposition period (2-day resolution), percent fertile females, total number of eggs per fertile female (after 24 days), daily fecundity (total number of eggs/number of survived days after the preoviposition period), and hatching rate (number of viable eggs/total number of eggs). The experiment was conducted twice with a total of 10–20 replicates per food treatment (Table 1).
Feeding experiments with S. littoralis larvae
In a second series of experiments, S. littoralis larvae were used as prey instead of spider mites. The experiments followed a similar protocol as described above with the following six food treatments (Table 1): SL: prey consisted of live larvae from either Bt or non-Bt cotton, presented on the Bt or non-Bt cotton leaf discs, respectively; SL/EE alt.: live larvae from Bt or non-Bt cotton were fed alternately with frozen E. kuehniella eggs in 2-day periods, while half of the replicates started with larvae as food and the other half with eggs; EE: prey consisted of E. kuehniella eggs, presented on Bt or non-Bt cotton leaf discs; EE/NF alt.: E. kuehniella eggs were fed alternately with no additional food in 2-day periods, while half of the replicates started with eggs and the other half with no additional food; NF: no additional food was provided on the Bt or non-Bt cotton leaf discs. SL + EE: live larvae from Bt or non-Bt cotton were fed simultaneously with E. kuehniella eggs. When leaf discs were changed (every second day), each O. majusculus pair received 10 new S. littoralis larvae in the respective treatments. Because of high consumption rates in the treatments with exclusive S. littoralis prey (SL), the number of larvae was increased to 15 from day 8 (first repetition) or day 6 (second repetition) onwards. Prey larvae still alive after the 2-day feeding periods were counted and numbers were subtracted from the number of prey larvae initially provided. In the following, this difference is referred to as “consumed prey”, although other causes of death or escapes might have contributed in addition to killing and consumption by the predators. The trials were terminated after 25 days (1 day after disc change). The experiment was conducted twice with a total of 10–20 replicates per food treatment (Table 1).
Bt protein analysis
During all experimental repetitions, samples of 7–20 mg fresh weight were collected from Bt and non-Bt cotton leaves (51 each), spider mites (11 Bt, 7 non-Bt), and S. littoralis (33 each) for the determination of Mpp51Aa2 using Enzyme-Linked Immunosorbent Assays (ELISA). Because the number of O. majusculus adults surviving the treatments with exclusive spider mite or S. littoralis feeding after 24 or 25 days was low, additional experiments were set up similar to the main experiments described above, but food treatments consisted exclusively of spider mites or S. littoralis larvae from either Bt or non-Bt cotton. Twenty individuals were set up for each food type. After 4 days (spider mites as food) or 5 days (larvae as food), all surviving O. majusculus were frozen and pooled to groups of 3–4 individuals. ELISA procedures followed the protocol described previously (Kim et al. 2021; Boss et al. 2023). Tissue was macerated in tris–borate buffer using a bead mill. Samples were diluted according to their expected Bt protein content (Bt cotton leaves 200–1000 × , Bt spider mites 200 × , Bt larvae 100 × , non-Bt spider mites 20 × , and Orius and other non-Bt samples undiluted). Ninety-six-well plates were coated with anti-Mpp51Aa2 mouse antibody and a goat anti-Mpp51Aa2 (IgG)-biotin construct served as detection antibody. TMB was added for the colour reaction, which was stopped after 10 min with 6 M phosphoric acid. Absorbance was read at 450 nm and concentrations were determined with an 8-point standard curve derived from purified Mpp51Aa2. Limits of detection were calculated based on 3 × SD of the optical densities (OD) of 4–6 blanks included on each plate.
Data analysis
Data, available in Online Resource 1, were analysed using R, version 3.6.3. (The R Foundation for Statistical Computing, Vienna, Austria). Survival of adult O. majusculus was analysed separately for males and females with the package “survival”. Other parameters were analysed with generalized linear models (GLM). Fixed factors were food type (F), cotton type (C), and their interaction (F × C). For F and C, contrasts were set to orthogonal. Analyses started with models including “experimental repetition” as a random factor, but the factor was removed as it explained no additional variation in most cases. For the distribution and link function used for each parameter, see Tables 2 and 3. Effects of factors and interactions were determined from ANOVA tables with Type III sum of squares (“car” package) with α = 0.05. Significant differences between individual food treatments were determined with Tukey tests (“emmeans” package). For differences in percent fecund females, Games Howell tests (“rstatix” package) were used, because of a lack of variance in some treatments. For survival and for parameters where F × C was significant, F was compared for each C type and C was compared for each F type separately (Tables 2 and 3).
Results
Feeding experiments with spider mites
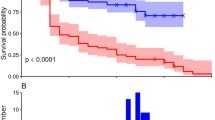
Longevity of O. majusculus females fed with spider mites reared on Bt cotton was 30% lower (14 days) than longevity of females fed with non-Bt mites (20 days) (p = 0.01, Table 2, Fig. 1A). By the end of the experiments (24 days), 4 of 20 females (20%) in the Bt cotton treatment and 10 of 20 females (50%) in the non-Bt cotton treatment were alive. When spider mites and E. kuehniella eggs were fed in alternation, 13 of 20 females (65%) survived the experimental period in the Bt and 17 of 20 females (85%) in the non-Bt group (p = 0.1). The different food treatments had a high impact on longevity (p < 0.0001). Without additional food (leaf disc only), O. majusculus females died on average after 5.7 days. Females fed with spider mites lived significantly longer than those without additional food. Females fed with E. kuehniella eggs, either exclusively or in alternation with spider mites or no additional food lived longest (Table 2). Male longevity showed a similar pattern (Table 2, Fig. 1B). Males fed spider mites from Bt lived half as long (6.2 days) as males fed spider mites from non-Bt cotton (12.7 days) (p = 0.0002). The different food types had a strong impact on male longevity (p < 0.0001).

Survival of Orius majusculus females (A) and males (B) raised on different food types: Tetranychus urticae spider mites (SM), Ephestia kuehniella eggs (EE), no additional food (NF), or two food types fed in alternation (SM/EE or EE/NF). Spider mites were reared either on Mpp51Aa2-producing cotton (Bt) or near-isogenic cotton (non-Bt). All prey types were presented on the respective Bt or non-Bt cotton leaf discs. Different letters indicate significant differences among food types (capital letters for non-Bt, small letters for Bt treatments). Asterisks indicate significant differences between cotton types (*p < 0.05, ***p < 0.001)
The percentage of fecund females was not influenced by cotton type (p = 1.0). Less fertile females were observed when only the leaf disc without additional food was provided to the predators compared with either E. kuehniella eggs as exclusive prey and eggs and no additional food in alternation (Table 2). No difference among cotton and food types was observed for the preoviposition time, which was on average 1.9–2.3 days (p ≥ 0.1, Table 2).
Total fecundity was less than half when O. majusculus females were fed with spider mites from Bt (10.5 eggs in total) compared with non-Bt cotton (27.7) (p < 0.0001, Fig. 2, Table 2). Similarly, significantly less eggs were laid when spider mites from Bt cotton and E. kuehniella eggs were fed in alternation (78.5) compared with the non-Bt treatment (115.0) (p = 0.01). Without additional food, females managed to lay on average 5.5 eggs, significantly less than when fed with spider mites. Compared with the spider mite treatments, significantly more eggs were produced when E. kuehniella eggs were provided, either exclusively or in alternation with spider mites or no additional food (Fig. 2, Table 2). A similar pattern was observed for daily fecundity. While spider mites as exclusive food resulted in fewer eggs in the Bt compared with the non-Bt treatment (p = 0.001), a comparable effect was borderline non-significant when spider mites and E. kuehniella eggs were fed in alternation (p = 0.06).

Total fecundity of Orius majusculus raised on different food types: Tetranychus urticae spider mites (SM), Ephestia kuehniella eggs (EE), no additional food (NF), or two food types fed in alternation (SM/EE alt. or EE/NF alt.). Spider mites were reared either on Mpp51Aa2-producing cotton (Bt) or near-isogenic cotton (non-Bt). All prey types were presented on the respective Bt or non-Bt cotton leaf discs. Dots represent individual values (N = 5–20), black rhombuses means, black horizontal lines medians, hinges 25th and 75th percentiles, and whiskers the smallest or largest values no further than 1.5 × IQR from the hinges. Results of GLM with fixed factors food (F) and cotton type (C) are presented in the grey box. Letters display sigificant differences between prey types, analysed separately for the two cotton types. Asterisks indicate significant differences between cotton types (*p < 0.05, **p < 0.01, ***p < 0.001)
The percentage of viable eggs (hatching rate) laid by O. majusculus females was not affected by cotton type in the treatments involving spider mites (p ≥ 0.06). However, the hatching rate of eggs from females fed with spider mites from Bt cotton was lower than the rate from females fed with E. kuehniella eggs, either exclusively or in alternation with spider mites or no additional food (Table 2).
For all parameters, no differences between the Bt and the non-Bt treatment were observed when no spider mites were involved as a vehicle for Mpp51Aa2 transfer. In those treatments, only the leaf disc contained the Bt protein, while E. kuehniella eggs, containing no Bt protein, were the same in both treatments. One exception was the hatching rate in exclusive E. kuehniella treatments, which was higher in the Bt compared with the non-Bt treatment (p = 0.001, Table 2).
Feeding experiments with S. littoralis larvae
In the experiments with S. littoralis larvae as prey, female longevity showed no significant differences between Bt and non-Bt treatments, except in the treatments without additional food, where females kept on Bt leaf discs lived 9.1 days and those on non-Bt leaf discs 7.2 days (Fig. 3A, Table 3). Females provided with food lived longer than in the treatments without additional food and females fed with S. littoralis larvae from Bt cotton exclusively had a higher mortality than females fed with Bt larvae and E. kuehniella eggs simultaneously (p < 0.05). Other food treatments had no impact on female longevity (Table 3, p ≥ 0.05). Orius majusculus males fed with S. littoralis larvae reared on Bt cotton survived half as long (8.7 days) as males fed with non-Bt larvae (15.4 days) (p = 0.01, Fig. 3B, Table 3). After 25 days, 1 of 19 males in the Bt cotton treatment and 6 of 20 males in the non-Bt cotton treatment were alive. In all treatments where E. kuehniella eggs were provided as food, more than 75% of the males survived the experimental period, regardless if Bt or non-Bt treatment. The different food types had a high impact on male longevity (p < 0.0001). Without additional food, O. majusculus males died on average after 4.4–4.8 days. Males fed with S. littoralis larvae lived significantly longer than those without additional food. Males fed with E. kuehniella eggs, either exclusively or in alternation with larvae or in alternation with no additional food, or simultaneously with larvae, lived longest (Fig. 3B, Table 3).

Survival of Orius majusculus females (A) and males (B) raised on different food types: Spodoptera littoralis larvae (SL), Ephestia kuehniella eggs (EE), no additional food (NF), two food types fed in alternation (SL/EE or EE/NF) or simultaneously (SL + EE). Larvae were fed with Mpp51Aa2-producing cotton (Bt) or near-isogenic cotton (non-Bt). All prey types were presented on the respective Bt or non-Bt cotton leaf discs. Different letters indicate significantly different food types (capital letters for non-Bt, small letters for Bt treatments). Asterisks indicate significant differences between cotton types (*p < 0.05)
The percentage of fecund females was not influenced by cotton type (p = 1.0) or food type (p = 0.2) (Table 3). While preoviposition time also was similar among cotton types (p = 0.9), females without additional food and females fed with E. kuehniella eggs exclusively produced first eggs earlier than females fed with S. littoralis larvae (exclusively or in alternation or simultaneously with E. kuehniella eggs). Longest preoviposition periods were observed for females fed with larvae exclusively.
Total fecundity was not affected by cotton type (p = 0.7, Fig. 4, Table 3), but food treatment had a strong effect (p < 0.0001). Without additional food, female O. majusculus managed to lay on average 5.6 eggs (non-Bt leaf discs) and 9.9 eggs (Bt leaf discs). More eggs were produced when females were provided S. littoralis larvae exclusively (38.5/33.6) and even more eggs when E. kuehniella eggs were fed, either in alternation, simultaneously, or exclusively (≥ 106). The highest fecundity was observed in the treatments with exclusive E. kuehniella food (170/185 eggs) (Fig. 4, Table 3). The same pattern was observed for daily fecundity (Table 3).

Total fecundity of Orius majusculus raised on different food types: Spodoptera littoralis larvae (SL), Ephestia kuehniella eggs (EE), no additional food (NF), two food types fed in alternation (SL/EE alt. or EE/NF alt.), or simultaneously (SL + EE). Larvae were reared either on Mpp51Aa2-producing cotton (Bt) or near-isogenic cotton (non-Bt). All prey types were presented on the respective Bt or non-Bt cotton leaf discs. Dots represent individual values (N = 8–20), black rhombuses means, black horizontal lines medians, hinges 25th and 75th percentiles, and whiskers the smallest or largest values no further than 1.5 × IQR from the hinges. Results of GLM with fixed factors food (F) and cotton type (C) are presented in the grey box (***p < 0.001). Letters display sigificant differences between prey types
The hatching rate of the eggs laid by O. majusculus females was not affected by cotton type (p = 0.5), while prey type was significant (p < 0.0001) (Table 3). The percentage of viable eggs was lower when the predators were fed S. littoralis larvae exclusively or when E. kuehniella eggs were alternated with no additional food, compared with the treatments with E. kuehniella eggs only or larvae + eggs simultaneously.
When analyzing the number of consumed prey larvae, a clear food-treatment effect was observed (p < 0.0001), while cotton type had no effect (p = 0.2). An average of 9.0–9.3 larvae was consumed (defined as not retrieved) per 2-day feeding period when larvae were the only available prey. In contrast, when larvae and E. kuehniella eggs were provided to the predators alternately, only 2.4 (Bt) or 3.4 (non-Bt) larvae were consumed. When eggs were available at the same time as larvae, approximately 1.1 larva was consumed after the 2-day feeding periods.
Mpp51Aa2 concentrations in leaves, prey and predators
Bt cotton leaves contained a median of 110 µg Mpp51Aa2/g fresh weight in the experiments with spider mites and 139 µg/g in the experiments with S. littoralis larvae (Table 4). Prey species contained 6–10 times less Bt protein compared to cotton leaves, i.e., spider mites 19.7 µg/g and larvae 15.0 µg/g. In O. majusculus feeding 4–5 days on Bt protein-containing prey, 100–500 times less Mpp51Aa2 was measured compared with their prey: 0.034 µg/g when fed with spider mites and 0.15 µg/g when fed with larvae (Table 4).
Discussion
Effects of Bt cotton on O. majusculus performance
When exclusively fed with frozen spider mites from Bt cotton, O. majusculus males and females showed reduced longevity and females produced fewer eggs (total fecundity and daily fecundity) compared with bugs fed with spider mites from non-Bt cotton. This confirms findings by Kim et al. (2021), who also reported reduced longevity and fecundity when O. majusculus females were fed with living spider mites from Mpp51Aa2-producing cotton. Because Kim et al. (2021) started their experiments with 5-day-old nymphs, their observed effects might have resulted from effects in the nymphal stage (leading to lower adult weight) in combination with effects during the adult feeding period. The current experiments, however, show clearly that Bt spider mites caused effects on male and female O. majusculus when test specimens had the same history in their developmental period until adulthood.
When O. majusculus adults were fed exclusively with S. littoralis larvae that have fed on Bt cotton, effects were only observed on male longevity, but not on female longevity or fecundity. Males suffered higher mortality compared with females in treatments without food or with food of suboptimal nutritional value, such as spider mites or S. littoralis larvae. This higher sensitivity of males to food quality also might have resulted in higher sensitivity towards the Bt toxin compared with females. Spodoptera littoralis larvae contained 25% less Mpp51Aa2 than spider mites. This rather small difference might be responsible for the observed difference in effects between prey types. However, it is likely that other factors, such as differences in metabolism that might have influenced the bioactivity of the measured Mpp51Aa2 in spider mites and larvae, as well as differences in the general suitability of the prey species for the predator might also have impacted the toxicity.
Similar to the current study on adults, Boss et al. (2023) reported no Bt effect when S. littoralis larvae were used as food for O. majusculus nymphs during their developmental period, but strong effects when spider mites were used. It has to be noted, however, that ELISA measurements suggested a larger difference in Mpp51Aa2 concentrations between spider mites (30–103 µg/g fresh weight) and larvae (18 µg/g) in the study by Boss et al. (2023) compared with the present study.
The key question of the current study was if supplementing a spider mite or S. littoralis diet with high quality food that does not contain Bt protein (E. kuehniella eggs) could mitigate Bt effects. This was realized by feeding O. majusculus with spider mites or larvae and E. kuehniella eggs in alternation using 2-day intervals. Predator longevity and fecundity was much better than when fed with spider mites or larvae alone. However, a significant Bt effect on total fecundity was still visible with spider mites as prey, although less pronounced than for the spider mite-only treatments. Once more, a similar effect was reported by Boss et al. (2023) on developmental parameters: feeding alternately mites and eggs reduced Bt effects on development time and adult weight, but did not completely compensate them (Boss et al. 2023). In the assay with S. littoralis as prey, the effect on male longevity was no longer observed when prey larvae were supplemented with E. kuehniella eggs and no Bt effects were evident when both prey types were provided simultaneously.
Effects of different food types on O. majusculus performance
This study confirms earlier findings that mites are of lower nutritional value than E. kuehniella eggs for O. majusculus (Boss et al. 2023; Kim et al. 2021) and other Orius species (Bonte et al. 2017). Nevertheless, the study shows that longevity and fecundity with mites were better than without additional food. This demonstrates that O. majusculus consumed the frozen spider mites in our experiment. Performance of adult bugs fed with S. littoralis larvae generally was better than that of O. majusculus fed with spider mites, although the current study does not allow a direct statistical comparison, because the two prey types were not fed in the same experiments. The numbers of consumed larvae clearly show that O. majusculus preyed readily on S. littoralis when no other food was available. Ephestia kuehniella eggs was the best food for predator survival and reproduction, also confirming previous results for this (Boss et al. 2023) and other Orius species (Bonte et al. 2017). Interestingly, when eggs and no additional food was supplied in alternation, performance of O. majusculus was only impaired to some degree and similar to the performance when eggs and mites or eggs and larvae were provided in alternation. In addition, the number of consumed larvae was low when fed in alternation with eggs and even lower when fed simultaneously with eggs. This shows that O. majusculus prefers more suitable (“higher quality”) food when available. Furthermore, the predator can cope well with 2-day gaps without additional food or without high quality food and consumes a larger quantity of lower quality food only when no high-quality food is available for longer time periods.
In the treatments with simultaneous availability of eggs and larvae, fecundity of O. majusculus females was somewhat lower (albeit not significant) than in the treatments where eggs were provided exclusively, which might be due to the fact that O. majusculus females might have been disturbed by the larvae. Also, preoviposition time was longer in treatments with larvae compared with treatments without larvae.
Suitability of frozen prey
Frozen spider mites dried out quickly, which likely reduced the ability or preference of O. majusculus to consume them. Thus, nutritional suitability of frozen mites might be considered lower than of living mites. To minimize this effect, new spider mites were provided daily. Compared with Kim et al. (2021) who fed live spider mites to O. majusculus until death (up to 5 weeks), daily fecundity in the current experiment was similar in the Bt treatment (0.9 eggs per day), but half in the non-Bt treatment (current study 1.47 eggs per day, Kim et al. 2.9 eggs per day). Nevertheless, frozen spider mites increased longevity and total fecundity of O. majusculus 2–4 times compared with no additional food. For E. kuehniella eggs, freezing is less of a problem as the egg shell remains intact and prevents drying out.
For the second set of experiments, living S. littoralis larvae were used because they were relatively easy to produce in suitable quantities. It was shown, however, that frozen Spodoptera sp. larvae also were suitable food for Orius sp. (Ren et al. 2022). Whenever necessary, freezing is thus a good option to store and ship large amounts of prey without the need for constant rearing infrastructure. However, suitability of frozen prey depends on prey type (e.g., eggs or larvae) and on the ability of the predator to handle and digest dead prey. In addition, Bt proteins are likely to deteriorate quicker in frozen and thawed prey items compared with living prey that can continuously feed on Bt plant leaf discs, which might have also affected the results of the current study.
Implications
Adverse effects of prey with low nutritional quality and high Mpp51Aa2 concentrations, such as spider mites, were evident for O. majusculus development (Boss et al. 2023), longevity, and reproduction (Kim et al. 2021 and this study) when representing the sole food source. Spider mites have been reported to contain the highest measured Bt protein concentrations among arthropods in Bt crops (Romeis et al. 2019; Torres and Ruberson 2008) and can thus be considered a worst-case exposure scenario for predators. Orius spp. have been reported to also feed on tissue of certain plants (Armer et al. 1998; Pumariño and Alomar 2012), which might expose them to even higher concentrations, if the respective plant produces insecticidal proteins. In the current leaf disc setup with cotton, however, the presence of plant material did not improve longevity compared with a treatment without leaf disc, suggesting that plant feeding in cotton is limited (Boss et al. 2023).
Alternative food can partly or fully mitigate Bt effects, depending on when and how long it is available. Orius spp. are known to consume a variety of soft-bodied insects in the field as well as pollen (Ballal and Yamada 2016; Corey et al. 1998; Lattin 1999). Whether or not the food scenarios deployed by Boss et al (2023) and in this study are realistic is difficult to judge. In the field, however, it is likely that a diverse mixture of prey species is available and that also the Bt protein concentrations of prey items range from zero to high (Eisenring et al. 2017). In particular phloem-feeding species, like aphids, whiteflies, and some leafhoppers contain no or only trace amounts of Bt protein (Eisenring et al. 2017; Meissle and Romeis 2018; Boss et al. 2023), as do non-feeding stages, such as insect eggs or pupae (Meissle et al. 2021). Furthermore, Mpp51Aa2 at doses produced in Bt cotton and observed in herbivorous prey species, resulted in significant, but limited effects on O. majusculus development, longevity and fecundity only in worst case exposure scenarios with spider mites (Kim et al. 2021; Boss et al. 2023, and this study). Choice experiments indicated, however, that spider mites are not a preferred prey of O. majusculus (Boss et al. 2023) and O. sauteri (Xu and Enkegaard 2009). When S. littoralis larvae were provided to O. majusculus as prey, no effects on female longevity and reproduction were evident despite relatively high Bt protein concentrations in the larvae (Boss et al. 2023 and this study). No effects were observed when spider mites and alternative prey were available simultaneously (Boss et al. 2023 and this study). The results of this study thus confirm the outcome of the environmental risk assessment (summarized in USDA-APHIS 2020) that predatory bug populations are unlikely to suffer harm from MON 88702 cotton in the field, because it is unlikely that their diet consists predominantly of spider mites and other prey types with similarly high Bt protein concentrations. In fact, arthropod communities have been studied in field experiments with MON 88702 cotton over three years at 5–6 sites in various cotton growing regions of the USA (Asiimwe et al. 2023). No significant differences in abundance of Orius spp. and other predatory Heteroptera in MON 88702 compared to unsprayed conventional cotton (DP393) were observed. As there is no evidence that biological control by predatory bugs is disrupted, MON 88702 cotton may thus be a valuable tool for the management of sucking pests in integrated production systems. Efficient control of target pests by MON 88702 in combination with functional populations of natural enemies may also contribute to a reduced need for pesticide applications, which is an important goal of integrated pest management.
Data availability
All data used for this manuscript are available in the electronic supplementary material.
References
Akbar W, Gowda A, Ahrens JE, Stelzer JW, Brown RS, Bollman SL, Greenplate JT, Gore J, Catchot AL, Lorenz G, Stewart SD, Kerns DL, Greene JK, Toews MD, Herbert DA, Reisig DD, Sword GA, Ellsworth PC, Godfrey LD, Clark TL (2019) First transgenic trait for control of plant bugs and thrips in cotton. Pest Manag Sci 75:867–877. https://doi.org/10.1002/ps.5234
Armer CA, Wiedenmann RN, Bush DR (1998) Plant feeding site selection on soybean by the facultatively phytophagous predator Orius insidiosus. Entomol Exp Appl 86:109–118. https://doi.org/10.1046/j.1570-7458.1998.00271.x
Asiimwe P, Brown CR, Ellsworth PC, Reisig DD, Bertho L, Jiang C, Schapaugh A, Head G, Burzio L (2023) Transgenic cotton expressing Mpp51Aa2 does not adversely impact beneficial non-target Hemiptera in the field. Crop Prot 173:106384. https://doi.org/10.1016/j.cropro.2023.106384
Bachman PM, Ahmad A, Ahrens JE, Akbar W, Baum JA, Brown S, Clark TL, Fridley JM, Gowda A, Greenplate JT, Jensen PD, Mueller GM, Odegaard ML, Tan J, Uffman JP, Levine SL (2017) Characterization of the activity spectrum of MON 88702 and the plant-incorporated protectant Cry51Aa2.834_16. PloS ONE 12:e0169409. https://doi.org/10.1371/journal.pone.0169409
Ballal CR, Yamada K (2016) Chapter 6 - Anthocorid Predators. In: Omkar (ed) Ecofriendly pest management for food security. Academic Press, San Diego, pp 183–216. https://doi.org/10.1016/B978-0-12-803265-7.00006-3
Bonte J, Van de Walle A, Conlong D, De Clercq P (2017) Eggs of Ephestia kuehniella and Ceratitis capitata, and motile stages of the astigmatid mites Tyrophagus putrescentiae and Carpoglyphus lactis as factitious foods for Orius spp. Insect Sci 24:613–622. https://doi.org/10.1111/1744-7917.12293
Boss A, Romeis J, Meissle M (2023) Prey-mediated effects of mCry51Aa2-producing cotton on the predatory nontarget bug Orius majusculus (Reuter). Insect Sci 30:1191–1206
Corey D, Kambhampati S, Wilde G (1998) Electrophoretic analysis of Orius insidiosus (Hemiptera: Anthocoridae) feeding habits in field corn. J Kansas Entomol Soc 71:11–17
Crickmore N, Berry C, Panneerselvam S, Mishra R, Connor TR, Bonning BC (2021) A structure-based nomenclature for Bacillus thuringiensis and other bacteria-derived pesticidal proteins. J Invertebr Pathol 186:107438. https://doi.org/10.1016/j.jip.2020.107438
Eisenring M, Romeis J, Naranjo SE, Meissle M (2017) Multitrophic Cry-protein flow in a dual-gene Bt-cotton field. Agric Ecosyst Environ 247:283–289. https://doi.org/10.1016/j.agee.2017.07.009
Ferragut F, González Zamora JE (1994) Diagnóstico y distribución de las especies de Orius Wolff 1811, peninsulares (Heteroptera, Anthocoridae). Bol San Veg Plagas 20:89–101
Gowda A, Rydel TJ, Wollacott AM, Brown RS, Akbar W, Clark TL, Flasinski S, Nageotte JR, Read AC, Shi X, Werner BJ, Pleau MJ, Baum JA (2016) A transgenic approach for controlling Lygus in cotton. Nat Commun 7:12213. https://doi.org/10.1038/ncomm12213
Kim Y, Kloos S, Romeis J, Meissle M (2021) Effects of mCry51Aa2-producing cotton on the non-target spider mite Tetranychus urticae and the predatory bug Orius majusculus. J Pest Sci 94:351–362. https://doi.org/10.1007/s10340-020-01260-4
Lattin JD (1999) Bionomics of the Anthocoridae. Annu Rev Entomol 44:207–231. https://doi.org/10.1146/annurev.ento.44.1.207
Meissle M, Romeis J (2018) Transfer of Cry1Ac and Cry2Ab proteins from genetically engineered Bt cotton to herbivores and predators. Insect Sci 25:823–832. https://doi.org/10.1111/1744-7917.12468
Meissle M, Kloos S, Romeis J (2021) Fate of multiple Bt proteins from stacked Bt maize in the predatory lady beetle Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae). Environ Pollut 268:115421. https://doi.org/10.1016/j.envpol.2020.115421
Meissle M, Grabenweger G, Romeis J (2023) No interaction of fluctuating or constant temperature and virulence of entomopathogenic fungi in two noctuid species. J Pest Sci. https://doi.org/10.1007/s10340-023-01673-x
Pumariño L, Alomar O (2012) The role of omnivory in the conservation of predators: Orius majusculus (Heteroptera: Anthocoridae) on sweet alyssum. Biol Control 62:24–28. https://doi.org/10.1016/j.biocontrol.2012.03.007
Ren X, Huang J, Li X, Zhang J, Zhang Z, Chen L, Hafeez M, Zhou S, Liu Y (2022) Frozen lepidopteran larvae as promising alternative factitious prey for rearing of Orius species. Entomol Gen 42:959–966. https://doi.org/10.1127/entomologia/2022/1579
Romeis J, Naranjo SE, Meissle M, Shelton AM (2019) Genetically engineered crops help support conservation biological control. Biol Control 130:136–154. https://doi.org/10.1016/j.biocontrol.2018.10.001
Torres JB, Ruberson JR (2008) Interactions of Bacillus thuringiensis Cry1Ac toxin in genetically engineered cotton with predatory heteropterans. Transgenic Res 17:345–354. https://doi.org/10.1007/s11248-007-9109-8
USDA-APHIS (2020) Monsanto petition (19-091-01p) for determination of non-regulated ststus of MON 88702 cotton. OECD Unique Identifier: MON-887Ø2–4. Final Plant Pest Risk Assessment. U.S. Department of Agriculture, Plant Health Inspection Service (USDA-APHIS). Available online: https://www.aphis.usda.gov/brs/aphisdocs/19-091-01p-ppra.pdf. Accessed 29 Aug 2023
Wijnands FG, Baur R, Malavolta C, Gerowitt B (2012) Integrated pest management—design and application of feasible and effective strategies. IOBC-WPRS, Lelystad
Xu X, Enkegaard A (2009) Prey preference of Orius sauteri between Western Flower Thrips and spider mites. Entomol Exp Appl 132:93–98. https://doi.org/10.1111/j.1570-7458.2009.00867.x
Acknowledgements
Bayer Crop Science (St. Louis, USA) is acknowledged for providing cotton seeds and ELISA reagents and Syngenta Crop Protection AG (Stein, Switzerland) for providing spider mites and noctuid larvae. I am grateful to Elina Leu, Mario Waldburger, and Tommy Hardeman for help in rearing predatory bugs and in performing bioassays. Further thanks go to Jörg Romeis (Agroscope) and Christopher Brown (Bayer) for valuable comments on an earlier draft of this manuscript.
Funding
Open access funding provided by Agroscope.
Author information
Authors and Affiliations
Contributions
The author confirms sole responsibility for the following: study conception and design, data collection, analysis and interpretation of results, and manuscript preparation.
Corresponding author
Ethics declarations
Conflict of interest
The author declares no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Meissle, M. Prey-mediated effects of Mpp51Aa2-producing cotton on longevity and reproduction of Orius majusculus. Transgenic Res (2024). https://doi.org/10.1007/s11248-024-00378-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11248-024-00378-w